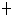 Gly bond in repressor LexA
Gly bond in repressor LexAEnzyme Nomenclature. Recommendations 1992
Prepared on behalf of the advisory panel on peptidase nomenclature by Alan J. Barrett, The Babraham Institute, Cambridge, England.
This supplement is as close as possible to the published version [see Eur. J. Biochem., 1995, 232, 1-6]. If you need to cite this supplement please quote this references as its source.
This document contains further additions and amendments to Enzyme Nomenclature 1992, published by Academic Press, Orlando, Florida. The previous Supplement was published in Eur. J. Biochem. 223, 1-5 (1994). Comments and suggestions on enzyme classification and nomenclature may be sent to Prof. K. F. Tipton, Department of Biochemistry, Trinity College, Dublin, Dublin 2, Ireland.
3.4.11.20 Aminopeptidase Ey
Reaction: Differs from other aminopeptidases in broad specificity for amino acids in the P1 position and the ability to hydrolyse peptides of four or five residues that contain Pro in the P1' position
Comments: A zinc glycoprotein comprised of two 150 kDa subunits. From the plasma fraction of hen egg yolk
References:
1. Ichishima, E., Yamagata, Y., Chiba, H., Sawaguchi, K. & Tanaka, T. (1989) Soluble and bound forms of aminopeptidase in hens egg-yolk. Agric. Biol. Chem. 53, 1867-1872
2. Tanaka, T. & Ichishima, E. (1993) Substrate specificity of aminopeptidase Ey from hen's egg yolk. Comp. Biochem. Physiol. [B] 105, 105-110
3. Tanaka, T. & Ichishima, E. (1993) Molecular properties of aminopeptidase Ey as a zinc-metalloenzyme. Int. J. Biochem. 25, 1681-1688
3.4.21.88 Repressor LexA
Reaction: Hydrolysis of Ala84Gly bond in repressor LexA
Other Names: LexA repressor
Comments: RecA protein and single-stranded DNA are required for activity, which is attributed to a Ser/Lys dyad [2]. The LexA protein represses the SOS regulon, which regulates the genes involved in DNA repair. In the presence of single-stranded DNA, the RecA protein interacts with repressor LexA, causing it to undergo an autocatalytic cleavage which disrupts the DNA-binding part of the repressor, and inactivates it. The consequent derepression of the SOS regulon leads to DNA repair. This peptidase activity of LexA was previously attributed to the RecA protein (formerly EC 3.4.99.37)
References:
1. Horii, T., Ogawa, T. & Ogawa, H. (1981) Nucleotide sequence of the LexA gene of E. coli. Cell 23, 689-697
2. Slilaty, S.N. & Little, J.W. (1987) Lysine-156 and serine-119 are required for LexA repressor cleavage: a possible mechanism. Proc. Natl. Acad. Sci. USA 84, 3987-3991
3. Kim, B. & Little, J.W. (1993) LexA and CI repressors as enzymes: specific cleavage in an intermolecular reaction. Cell 73, 1165-1173
4. Little, J.W., Kim, B., Roland, K.L., Smith, M.H., Lin, L.-L. & Slilaty, S.N. (1994) Cleavage of LexA repressor. Methods Enzymol. 244, 266-284
3.4.21.89 Signal peptidase I
Reaction: Cleavage of hydrophobic, N-terminal signal or leader sequences from secreted and periplasmic proteins
Other Names: Leader peptidase I
Comments: Present in the inner membrane of E. coli. Unaffected by inhibitors of most serine peptidases, but site-directed mutagenesis implicates a Ser/Lys catalytic dyad in activity [1,3]. Hydrolyses a single bond -AlaAla- in M13 phage procoat protein, producing free signal peptide and coat protein. Formerly included in EC 3.4.99.36. Eukaryote signal peptidases that may have somewhat different specificity are known from the endoplasmic reticulum membrane [4] and mitochondrial inner membrane [2]
References:
1. Black, M.T. (1993) Evidence that the catalytic activity of prokaryote leader peptidase depends upon the operation of a serine-lysine catalytic dyad. J. Bacteriol. 175, 4957-4961
2. Nunnari, J., Fox, T.D. & Walter, P. (1993) A mitochondrial protease with two catalytic subunits of nonoverlapping specificities. Science 262, 1997-2004
3. Tschantz, W.R., Sung, M., Delgado-Partin, V.M. & Dalbey, R.E. (1993) A serine and a lysine residue implicated in the catalytic mechanism of the Escherichia coli leader peptidase. J. Biol. Chem. 268, 27349-27354
4. Lively, M.O., Newsome, A.L. & Nusier, M. (1994) Eukaryote microsomal signal peptidases. Methods Enzymol. 244, 301-314
5. Tschantz, W.R. & Dalbey, R.E. (1994) Bacterial leader peptidase I. Methods Enzymol. 244, 285-301
3.4.21.90 Togavirin
Reaction: Autocatalytic release of the core protein from the N-terminus of the togavirus structural polyprotein by hydrolysis of a -TrpSer- bond
Other Names: Sindbis virus protease; Sindbis virus core protein; nsP2 proteinase
Comments: Known from the Sindbis and Semliki forest togaviruses. Once released, the core protein does not retain catalytic activity. Togavirin has a similar tertiary structure to chymotrypsin [3]
References:
1. Kräusslich, H.-G. & Wimmer, E. (1988) Viral proteinases. Annu. Rev. Biochem. 57, 701-754
2. Strauss, E.G., De Groot, R.J., Levinson, R. & Strauss, J.H. (1992) Identification of the active site residues in the nsP2 proteinase of Sindbis virus. Virology 191, 932-940
3. Tong, L., Wengler, G. & Rossmann, M.G. (1993) Refined structure of Sindbis virus core protein and comparison with other chymotrypsin-like serine proteinase structures. J. Mol. Biol. 230, 228-247
3.4.21.91 Flavivirin
Reaction: Selective hydrolysis of -Xaa-XaaXbb- bonds in which each of the Xaa can be either Arg or Lys and Xbb can be either Ser or Ala
Other Names: Yellow fever virus (flavivirus) protease; NS2B-3 proteinase
Comments: Known from classical flaviviruses (yellow fever, dengue fever). The functional viral peptidase is part of the NS2B protein. Catalytic His, Asp and Ser residues are arranged as in chymotrypsin
References:
1. Chambers, T.J., Hahn, C.S., Galler, R. & Rice, C.M. (1990) Flavivirus genome organization, expression, and replication. Annu. Rev. Microbiol. 44, 649-688
2. Cahour, A., Falgout, B. & Lai, C.-J. (1992) Cleavage of the dengue virus polyprotein at the NS3/NS4A and NS4B/NS5 junctions is mediated by viral protease NS2B-NS3, whereas NS4A/NS4B may be processed by a cellular protease. J. Virol. 66, 1535-1542
3. Lin, C., Amberg, S.M., Chambers, T.J. & Rice, C.M. (1993) Cleavage at a novel site in the NS4A region by the yellow fever virus NS2B-3 proteinase is a prerequisite for processing at the downstream 4A/4B signalase site. J. Virol. 67, 2327-2335
3.4.23.36 Signal peptidase II
Reaction: Release of signal peptides from bacterial membrane prolipoproteins including murein prolipoprotein. Hydrolyses -Xaa-Xbb-Xcc(S,diacylglyceryl)Cys-, in which Xaa is hydrophobic (preferably Leu), and Xbb (Ala or Ser) and Xcc (Gly or Ala) have small, neutral sidechains
Other Names: Premurein-leader peptidase; Prolipoprotein signal peptidase; Leader peptidase II
Comments: An 18-kDa enzyme present in bacterial inner membranes. Inhibited by pepstatin and the antibiotic globomycin. Formerly EC 3.4.99.35
References:
1. Dev, I.K. & Ray, P.H. (1990) Signal peptidases and signal peptide hydrolases. J. Bioenerg. Biomembr. 22, 271-290
2. Zhao, X.-J. & Wu, H.C. (1992) Nucleotide sequence of the Staphylococcus aureus signal peptidase II (lsp) gene. FEBS Lett. 299, 80-84
3. Sankaran, K. & Wu, H.C. (1994) Bacterial prolipoprotein signal peptidase. Methods Enzymol. 248, 169-180
3.4.23.37 Pseudomonapepsin
Reaction: Hydrolysis of the B chain of insulin at -Glu13Ala-, -Leu15Tyr- and -Phe25Tyr-, and angiotensin I at -Tyr4Ile-. A good synthetic substrate is Lys-Pro-Ile-Glu-PhePhe(NO2)-Arg-Leu.
Other Names: Pseudomonas sp. pepstatin-insensitive carboxyl proteinase
Comments: An enzyme secreted by Pseudomonas sp. No. 101 that is insensitive to pepstatin, diazoacetylnorleucine methyl ester and 1,2-epoxy-3-(p-nitrophenoxy)propane (EPNP). Optimum pH 4. It is distinguished from xanthomonapepsin by insensitivity to EPNP and from scytalidopepsin B by this property and by its unrelated amino acid sequence. Inhibited by tyrostatin, a peptide aldehyde [2]
References:
1. Oda, K., Sugitani, M., Fukuhara, K. & Murao, S. (1987) Purification and properties of a pepstatin-insensitive carboxyl proteinase from a Gram-negative bacterium. Biochim. Biophys. Acta 923, 463-469
2. Oda, K., Fukuda, Y., Murao, S., Uchida, K. & Kainosho, M. (1989) A novel proteinase inhibitor, tyrostatin, inhibiting some pepstatin-insensitive carboxyl proteinases. Agric. Biol. Chem. 53, 405-415
3. Oda, K. & Murao, S. (1991) Pepstatin-insensitive carboxyl proteinases. In Structure and Function of Aspartic Proteinases (Dunn, B.M., ed.), pp. 185-201, Plenum Press, New York
4. Oda, K., Nakatani, H. & Dunn, B.M. (1992) Substrate specificity and kinetic properties of pepstatin-insensitive carboxyl proteinase from Pseudomonas sp. No. 101. Biochim. Biophys. Acta 1120, 208-214
3.4.23.38 Plasmepsin I
Reaction: Hydrolysis of the -Phe3+Leu- bond in the α-chain of hemoglobin, leading to denaturation of the molecule
Other Names: Aspartic hemoglobinase I; PFAPG
Comments: Known from the malaria organism, Plasmodium. About 37 kDa. Member of the pepsin family, closest to cathepsin D and renin in structure. Inhibited by pepstatin. Formerly included in EC 3.4.23.6
References:
1. Goldberg, D.E., Slater, A.F.G., Beavis, R., Chait, B., Cerami, A. & Henderson, G.B. (1991) Hemoglobin degradation in the human malaria pathogen Plasmodium falciparum: a catabolic pathway initiated by a specific aspartic protease. J. Exp. Med. 173, 961-969.
2. Francis, S.E., Gluzman, I.Y., Oksman, A., Knickerbocker, A., Mueller, R., Bryant, M.L., Sherman, D.R., Russell, D.G. & Goldberg, D.E. (1994) Molecular characterization and inhibition of a Plasmodium falciparum aspartic hemoglobinase. EMBO J. 13, 306-317
3. Gluzman, I.Y., Francis, S.E., Oksman, A., Smith, C.E., Duffin, K.L. & Goldberg, D.E. (1994) Order and specificity of the Plasmodium falciparum hemoglobin degradation pathway. J. Clin. Invest. 93, 1602-1608
3.4.23.39 Plasmepsin II
Reaction: Hydrolysis of the bonds linking certain hydrophobic residues in hemoglobin or globin. Also cleaves the small molecule substrates such as Ala-Leu-Glu-Arg-Thr-PhePhe(NO2)-Ser-Phe-Pro-Thr [3]
Other Names: Aspartic hemoglobinase II; PFAPD
Comments: Known from the malaria organism, Plasmodium. About 37 kDa. Member of the pepsin family, 73% identical in sequence to plasmepsin I. Inhibited by pepstatin. Formerly included in EC 3.4.23.6
References:
1. Dame, J.B., Reddy, G.R., Yowell, C.A., Dunn, B.M., Kay, J. & Berry, C. (1994) Sequence, expression and modelled structure of an aspartic proteinase from the human malaria parasite Plasmodium falciparum. Mol. Biochem. Parasitol. 64, 177-190
2. Gluzman, I.Y., Francis, S.E., Oksman, A., Smith, C.E., Duffin, K.L. & Goldberg, D.E. (1994) Order and specificity of the Plasmodium falciparum hemoglobin degradation pathway. J. Clin. Invest. 93, 1602-1608
3. Hill, J., Tyas, L., Phylip, L.H., Kay, J., Dunn, B.M. & Berry, C. (1994) High level expression and characterisation of plasmepsin II, an aspartic proteinase from Plasmodium falciparum. FEBS Lett. 352, 155-158
3.4.24.60 Dactylysin
Reaction: Hydrolysis of peptides of at least six residues, with bulky hydrophobic residues in the P1 position. Shows a preference for hydrophobic doublets such as -PhePhe- and -PheLeu- in somatostatin-(1-14)-peptide and dynorphin A-(1-6)-peptide, respectively
Other Names: Peptide hormone inactivating endopeptidase
Comments: An endopeptidase of 100 kDa secreted from the skin of the amphibian, Xenopus laevis (Dactylêtre du Cap). Resembles neprilysin in insensitivity to 1 μM captopril, but differs from it in being insensitive to thiorphan (1 μM) and unable to digest [Met5]enkephalin, [Leu5]enkephalin, oxytocin, and substance P-(7-11)-peptide. A similar endopeptidase is found in human neuroblastoma cells [2]
References:
1. Carvalho, K.M., Joudiou, C., Boussetta, H., Leseney, A.-M. & Cohen, P. (1992) A peptide-hormone-inactivating endopeptidase in Xenopus laevis skin secretion. Proc. Natl. Acad. Sci. USA 89, 84-88
2. Delporte, C., Carvalho, K.M., Leseney, A.-M., Winand, J., Christophe, J. and Cohen, P. (1992) A new metallo-endopeptidase from human neuroblastoma NB-OK-1 cells which inactivates atrial natriuretic peptide by selective cleavage at the Ser123-Phe124 bond. Biochem. Biophys. Res. Commun. 182, 158-164
3. Joudiou, C., Carvalho, K.M., Camarao, G., Boussetta, H. & Cohen, P. (1993) Characterization of the thermolysin-like cleavage of biologically active peptides by Xenopus laevis peptide hormone inactivating enzyme. Biochemistry 32, 5959-5966
3.4.24.61 Nardilysin
Reaction: Hydrolysis of polypeptides, preferably at -XaaArg-Lys-, and less commonly at -ArgArg-Xaa-, in which Xaa is not Arg or Lys
Other Names: N-Arginine dibasic convertase; NRD-convertase
Comments: Enzyme of 133 kDa from rat brain and testis. A homologue of pitrilysin containing the His-Phe-Leu-Glu-His zinc-binding sequence, and a highly acidic stretch of 71 residues. Unusually for a metalloendopeptidase, inhibited by bestatin, amastatin and N-ethylmaleimide
References:
1. Gomez, S., Gluschankof, P., Morel, A. & Cohen, P. (1985) The somatostatin-28 convertase of rat brain cortex is associated with secretory granule membranes. J. Biol. Chem. 260, 10541-10545
2. Gluschankof, P., Gomez, S., Morel, A. & Cohen, P. (1987) Enzymes that process somatostatin precursors. A novel endoprotease that cleaves before the arginine-lysine doublet is involved in somatostatin-28 convertase activity of rat brain cortex. J. Biol. Chem. 262, 9615-9620
3. Chesneau, V., Pierotti, A.R., Barré, N., Créminon, C., Tougard, C. & Cohen, P. (1994) Isolation and characterization of a dibasic selective metalloendopeptidase from rat testes that cleaves at the amino terminus of arginine residues. J. Biol. Chem. 269, 2056-2061
4. Pierotti, A.R., Prat, A., Chesneau, V., Gaudoux, F., Leseney, A.-M., Foulon, T. & Cohen, P. (1994) N-arginine dibasic convertase, a metalloendopeptidase as a prototype of a class of processing enzymes. Proc. Natl. Acad. Sci. USA 91, 6078-6082
3.4.24.62 Magnolysin
Reaction: Hydrolysis of polypeptides with Arg or Lys in P1 and P2, e.g. to hydrolyse pro-oxytocin at -Lys-ArgAla-Val-. The specificity further depends on the organization of a β-turn-α-helix of nine or more residues containing the paired basic amino acids near the centre [3]
Other Names: Bovine neurosecretory granule protease cleaving pro-oxytocin/neurophysin
Comments: An endopeptidase of 58 kDa known from bovine pituitary neurosecretory granules and bovine and human corpus luteum [4,5]. Inhibited by EDTA [1]
References:
1. Clamagirand, C., Creminon, C., Fahy, C., Boussetta, H. & Cohen, P. (1987) Partial purification and functional properties of an endoprotease from bovine neurosecretory granules cleaving proocytosin/neurophysin peptides at the basic amino acid doublet. Biochemistry 26, 6018-6023
2. Créminon, C., Rholam, M., Boussetta, H., Marrakchi, N. & Cohen, P. (1988) Synthetic peptide substrates as models to study a pro-ocytocin/neurophysin converting enzyme. J. Chromatogt. 440, 439-448.
3. Brakch, N., Boussetta, H., Rholam, M. & Cohen, P. (1989) Processing endoprotease recognizes a structural feature at the cleavage site of peptide prohormones. The pro-ocytocin/neurophysin model. J. Biol. Chem. 264, 15912-15916.
4. Plevrakis, I, Créminon, C., Clamagirand, C., Brakch, N. Rholam, M. & Cohen, P. (1989) Proocytocin/neurophysin convertase from bovine neurohypophysis and corpus luteum secretory granules: complete purification, structure-function relationships, and competitive inhibitor. Biochemistry 28, 2705-2710.
5. Guillou, M.D., Camier, M. & Clamagirand, C. (1994) Evidence for the presence of pro-oxytocin/neurophysin converting enzyme in the human ovary. J. Endocrinol. 142, 345-352
3.4.24.63 Meprin B
Reaction: Hydrolysis of proteins, including azocasein, and peptides. Hydrolysis of -His5Leu-, -Leu6Cys-, -Ala14Leu- and -Cys19Gly- bonds in insulin B chain
Other Names: Meprin-b
Comments: A brush border membrane-bound metalloendopeptidase known from the intestine of all mouse strains that have been tested, and the kidney of certain inbred strains. A tetramer of meprin subunits (in contrast to meprin A, which contains both α and β subunits). Occurs in the kidney as a proenzyme that can be activated by trypsin. Meprin B is inhibited by both EDTA and 1,10-phenanthroline, but not by phosphoramidon, captopril or thiorphan. A homologue of astacin
References:
1. Kounnas, M.Z., Wolz, R.L., Gorbea, C.M. & Bond, J.S. (1991) Meprin-A and -B. Cell surface endopeptidases of the mouse kidney. J. Biol. Chem. 266, 17350-17357
2. Gorbea, C.M., Marchand, P., Jiang, W., Copeland, N.G., Gilbert, D.J., Jenkins, N.A. & Bond, J.S. (1993) Cloning, expression, and chromosomal localization of the mouse meprin subunit. J. Biol. Chem. 268, 21035-21043
3. Johnson, G.D. & Hersh, L.B. (1994) Expression of meprin subunit precursors. Membrane anchoring through the subunit and mechanism of zymogen activation. J. Biol. Chem. 269, 7682-7688
4. Wolz, R.L. & Bond, J.S. (1995) Meprins A and B. Methods Enzymol. 248, 325-345
3.4.24.64 Mitochondrial processing peptidase
Reaction: Release of N-terminal targetting peptides from precursor proteins imported into the mitochondrion, typically with Arg in position P2
Other Names: Processing enhancing peptidase (for one of two subunits)
Comments: Known from the mitochondrial matrix of fungi and mammals. Formed from two subunits, both homologous with pitrilysin [3], and the products of the MAS1 and MAS2 genes in yeast. Formerly EC 3.4.99.41
References:
1. Jensen, R.E. & Yaffe, M.P. (1988) Import of proteins into yeast mitochondria: the nuclear MAS2 gene encodes a component of the processing protease that is homologous to the MAS1-encoded subunit. EMBO J. 7, 3863-3871
2. Witte, C., Jensen, R.E., Yaffe, M.P. & Schatz, G. (1988) MAS1, a gene essential for yeast mitochondrial assembly, encodes a subunit of the mitochondrial processing protease. EMBO J. 7, 1439-1447
3. Rawlings, N.D. & Barrett, A.J. (1991) Homologues of insulinase, a new superfamily of metalloendopeptidases. Biochem. J. 275, 389-391
4. Kalousek, F., Neupert, W., Omura, T., Schatz, G. & Schmitz, U.K. (1993) Uniform nomenclature for the mitochondrial peptidases cleaving precursors of mitochondrial proteins. Trends Biochem. Sci. 18, 249 only
5. Brunner, M. & Neupert, W. (1994) Purification and characterization of the mitochondrial processing peptidase of Neurospora crassa. Methods Enzymol. 248, 717-728
3.4.24.65 Macrophage elastase
Reaction: Hydrolysis of soluble and insoluble elastin [1]. Specific cleavages are also produced at -Ala14Leu- and -Tyr16Leu- in the B chain of insulin [2]
Other Names: Metalloelastase
Comments: This enzyme is synthesized as a proenzyme of 53 kDa that is converted to an active form of 22 kDa. cDNA sequences have been obtained for the mouse [3] and human [4] enzymes. Homologue of interstitial collagenase
References:
1. Banda, M.J. & Werb, Z. (1981) Mouse macrophage elastase. Purification and characterization as a metalloproteinase. Biochem. J. 193, 589-605
2. Kettner, C., Shaw, E., White, R. & Janoff, A. (1981) The specificity of macrophage elastase on the insulin B-chain. Biochem. J. 195, 369-372
3. Shapiro, S.D., Griffin, G.L., Gilbert, D.J., Jenkins, N.A., Copeland, N.G., Welgus, H.G., Senior, R.M. & Ley, T.J. (1992) Molecular cloning, chromosomal localization, and bacterial expression of a murine macrophage metalloelastase. J. Biol. Chem. 267, 4664-4671
4. Shapiro, S.D., Kobayashi, D.K. & Ley, T.J. (1993) Cloning and characterization of a unique elastolytic metalloproteinase produced by human alveolar macrophages. J. Biol. Chem. 268, 23824-23829
3.4.24.66 Choriolysin L
Reaction: Hydrolysis of the inner layer of fish egg envelope. Also hydrolysis of casein and small molecule substrates such as succinyl-Leu-Leu-Val-Tyr7-(4-methyl)coumarylamide
Other Names: Teleost hatching enzyme (component); Low choriolytic enzyme (LCE)
Comments: Known from the teleost fish Oryzias latipes (medaka). Efficient dissolution of the egg membrane requires concerted action with choriolysin H. 24 kDa homologue of astacin
References:
1. Yasumasu, S., Iuchi, I. & Yamagami, K. (1988) Medaka hatching enzyme consists of two kinds of proteases which act cooperatively. Zool. Sci. 5, 191-195
2. Yasumasu, S., Iuchi, I. & Yamagami, K. (1989) Isolation and some properties of low choriolytic enzyme (LCE), a component of the hatching enzyme of the teleost, Oryzias latipes. J. Biochem. (Tokyo) 105, 212-218
3. Yasumasu, S., Katow, S., Hamazaki, T.S., Iuchi, I. & Yamagami, K. (1992) Two constituent proteases of a teleostean hatching enzyme: concurrent syntheses and packaging in the same secretory granules in discrete arrangement. Dev. Biol. 149, 349-356
4. Yasumasu, S., Yamada, K., Akasaka, K., Mitsunaga, K., Iuchi, I., Shimada, H. & Yamagami, K. (1992) Isolation of cDNAs for LCE and HCE, two constituent proteases of the hatching enzyme of Oryzias latipes, and concurrent expression of their mRNAs during development. Dev. Biol. 153, 250-258
3.4.24.67 Choriolysin H
Reaction: Hydrolysis of the inner layer of fish egg envelope. Also hydrolysis of casein and small molecule substrates such as succinyl-Leu-Leu-Val-Tyr7-(4-methyl)coumarylamide
Other Names: Teleost hatching enzyme (component); High choriolytic enzyme (HCE)
Comments: Known from the teleost fish Oryzias latipes (medaka). Efficient dissolution of the egg membrane requires concerted action with choriolysin L. 25.5 kDa homologue of astacin
References:
1. Yamagami, K. (1972) Isolation of a choriolytic enzyme (hatching enzyme) of the teleost, Oryzias latipes. Dev. Biol. 29, 343-348
2. Yasumasu, S., Iuchi, I. & Yamagami, K. (1989) Purification and partial characterization of high choriolytic enzyme (HCE), a component of the hatching enzyme of the teleost, Oryzias latipes. J. Biochem. (Tokyo) 105, 204-211
3. Yasumasu, S., Katow, S., Umino, Y., Iuchi, I. & Yamagami, K. (1989) A unique proteolytic action of HCE, a constituent protease of a fish hatching enzyme: tight binding to its natural substrate, egg envelope. Biochem. Biophys. Res. Commun. 162, 58-63
4. Yasumasu, S., Yamada, K., Akasaka, K., Mitsunaga, K., Iuchi, I., Shimada, H. & Yamagami, K. (1992) Isolation of cDNAs for LCE and HCE, two constituent proteases of the hatching enzyme of Oryzias latipes, and concurrent expression of their mRNAs during development. Dev. Biol. 153, 250-258
5. Lee, K.S., Yasumasu, S., Nomura, K. & Iuchi, I. (1993) HCE, a constituent of the hatching enzymes of Oryzias latipes embryos, releases unique proline-rich polypeptides from its natural substrate, the hardened chorion. FEBS Lett. 339, 281-284
3.4.24.68 Tentoxilysin
Reaction: Hydrolysis of -Gln76Phe- bond in synaptobrevin (also known as neuronal vesicle-associated membrane protein, VAMP)
Other Names: Tetanus neurotoxin
Comments: Zinc enzyme produced by Clostridium tetani. Proenzyme of 150 kDa is processed to disulfide-linked subunits of 100 and 50 kDa, the latter being responsible for the endopeptidase activity. Weakly inhibited by captopril, and phosphoramidon. The clostridial neurotoxins disable the neuroexocytosis apparatus, and have been described as the most toxic substances known. Tentoxilysin acts at the spinal inhibitory interneurons, blocking the release of various neurotransmitters to produce spastic paralysis
References:
1. Fujii, N., Kimura, K., Yashiki, T., Tsuzuki, K., Moriishi, K., Yokosawa, N., Syuto, B. & Oguma, K. (1992) A zinc-protease specific domain in botulinum and tetanus neurotoxins. Microbiol. Intern. 36, 213-220
2. Schiavo, G., Benfenati, F., Poulain, B., Rossetto, O., Polverino de Laureto, P., DasGupta, B.R. & Montecucco, C. (1992) Tetanus and botulinum-B neurotoxins block neurotransmitter release by proteolytic cleavage of synaptobrevin. Nature 359, 832-834
3. Schiavo, G., Rossetto, O., Santucci, A., DasGupta, B.R. & Montecucco, C. (1992) Botulinum neurotoxins are zinc proteins. J. Biol. Chem. 267, 23479-23483
4. Montecucco, C. & Schiavo, G. (1994) Mechanism of action of tetanus and botulinum neurotoxins. Mol. Microbiol. 8, 1-13
5. Montecucco, C. & Schiavo, G. (1995) Clostridial toxins. Methods Enzymol. 248, 643-652
3.4.24.69 Bontoxilysin
Reaction: Limited hydrolysis of proteins of the neuroexocytosis apparatus, synaptobrevin (also known as neuronal vesicle-associated membrane protein, VAMP), synaptosome-associated protein of 25 kDa (SNAP25) or syntaxin. No detected action on small molecule substrates
Other Names: Botulinum neurotoxin
Comments: This zinc enzyme, produced by Clostridium botulinum, occurs as forms A-G that differ in specificity of action on the proteins of the neuroexocytosis apparatus [1-5]. The 150-kDa proenzymes of bontoxilysin are processed to disulfide-linked subunits of 100 and 50 kDa, the latter being responsible for the endopeptidase activities. Weakly inhibited by captopril, and phosphoramidon. Toxicity is due to action at the neuromuscular junctions that blocks release of acetylcholine, causing flaccid paralysis, in contrast to the spastic paralysis caused by tentoxilysin
References:
1. Schiavo, G., Rossetto, O., Catsicas, S., Polverino De Laureto, P., DasGupta, B.R., Benfenati, F. & Montecucco, C. (1993) Identification of the nerve-terminal targets of botulinum neurotoxins serotypes A, D and E. J. Biol. Chem. 268, 23784-23787
2. Schiavo, G., Santucci, A., DasGupta, B.R., Mehta, P.P., Jontes, J., Benfenati, F., Wilson, M.C. & Montecucco, C. (1993) Botulinum neurotoxins serotypes A and E cleave SNAP-25 at distinct COOH-terminal peptide bonds. FEBS Lett. 335, 99-103
3. Schiavo, G., Shone, C.C., Rossetto, O., Alexander, F.C.G. & Montecucco, C. (1993) Botulinum neurotoxin serotype F is a zinc endopeptidase specific for VAMP/synaptobrevin. J. Biol. Chem. 268, 11516-11519.
4. Schiavo, G., Malizio, C., Trimble, W.S., Polverino De Laureto, P., Milan, G., Sugiyama, H., Johnson, E.A. & Montecucco, C. (1994) Botulinum G neurotoxin cleaves VAMP/synaptobrevin at a single Ala-Ala peptide bond. J. Biol. Chem. 269, 20213-20216
5. Montecucco, C. & Schiavo, G. (1994) Mechanism of action of tetanus and botulinum neurotoxins. Mol. Microbiol. 8, 1-13
6. Montecucco, C. & Schiavo, G. (1995) Clostridial toxins. Methods Enzymol. 248, 643-652
[3.4.99.35 Transferred entry: now EC 3.4.23.36 - Signal peptidase II]
[3.4.99.36 Transferred entry: now EC 3.4.21.89 - Signal peptidase I]
[3.4.99.41 Transferred entry: now EC 3.4.24.64 - Mitochondrial processing peptidase]
Return to Enzymes Nomenclature home page
Return to main IUBMB Biochemical Nomenclature home page