Recommendations for nomenclature and tables in biochemical thermodynamics
(Recommendation 1994)
5 Thermodynamic Background
Continued from 4.4 Experimental matters
Contents of Section
5 Thermodynamic Background
5.1 Transformed thermodynamic properties
5.2 Isomer group thermodynamics
5.3 Adjustment for ionic strength
5.4 Adjustment of standard thermodynamic properties of species to the desired pH and pMg
5.5 Calculation of the standard formation properties of a pseudoisomer group at specified pH and pMg
5.6 The actual experiment and thought experiments
5.7 Linear algebra
References for this Section
5. THERMODYNAMIC BACKGROUND
5.1 TRANSFORMED THERMODYNAMIC PROPERTIES
The definition of a transformed Gibbs energy is a continuation of a process that starts with the first and second laws of thermodynamics, but is not always discussed in terms of Legendre transforms. The combined first and second law for a closed system involving only pressure-volume work is
dU = TdS - PdV . . . . . . . . (45)
where U is the internal energy and S is the entropy. The criterion for spontaneous change at specified S and V is (dU )S,V ![[less than or equal]](../../greek/LTOREQ.GIF) 0. That is, if S and V are held constant, U can only decrease and is at a minimum at equilibrium. To obtain a criterion at specified S and P, the enthalpy was defined by the Legendre transform H = U + PV so that (dH )S,P 0. To obtain a criterion at specifed T and V, the Helmholtz energy was defined by the Legendre transform A = U - TS so that (dA )T,V 0. To obtain a criterion at specified T and P, the Gibbs energy was defined by the Legendre transform G = H - TS so that (dG )T,P 0. The Gibbs energy is especially useful because it provides the criterion for equilibrium at specified T and P. Two Legendre transforms can be combined. For example, the internal energy can be transformed directly to G by use of G = U + PV - TS. Alberty and Oppenheim (ref. 20, 21) used a Legendre transform to develop a criterion for equilibrium for the alkylation of benzene by ethylene at a specified partial pressure of ethylene. Wyman and Gill (ref. 22) have described the use of transformed Gibbs energies in describing macromolecular components in solution.
0. That is, if S and V are held constant, U can only decrease and is at a minimum at equilibrium. To obtain a criterion at specified S and P, the enthalpy was defined by the Legendre transform H = U + PV so that (dH )S,P 0. To obtain a criterion at specifed T and V, the Helmholtz energy was defined by the Legendre transform A = U - TS so that (dA )T,V 0. To obtain a criterion at specified T and P, the Gibbs energy was defined by the Legendre transform G = H - TS so that (dG )T,P 0. The Gibbs energy is especially useful because it provides the criterion for equilibrium at specified T and P. Two Legendre transforms can be combined. For example, the internal energy can be transformed directly to G by use of G = U + PV - TS. Alberty and Oppenheim (ref. 20, 21) used a Legendre transform to develop a criterion for equilibrium for the alkylation of benzene by ethylene at a specified partial pressure of ethylene. Wyman and Gill (ref. 22) have described the use of transformed Gibbs energies in describing macromolecular components in solution.
In 1992, Alberty (refs. 4, 5) used the Legendre transform
G ' = G - n '(H+)![[mu]](../../greek/MU.GIF) (H+) - n '(Mg2+)(Mg2+) . . . . . . . . (46)
(H+) - n '(Mg2+)(Mg2+) . . . . . . . . (46)
to define a transformed Gibbs energy G ' in terms of the Gibbs energy G. Here n '(H+) is the total amount of H+ in the system (bound and unbound) and (H+) is the specified chemical potential for H+, which is given for an ideal solution by
(H+) = (H+)o + RT ln ([H+]/c o) . . . . . . . . (47)
where (H+)o is the chemical potential of H+ at 1 M in an ideal solution at specified T, P, and I. The transformed Gibbs energy G ' is defined in order to obtain a criterion of spontaneous change at T, P, pH, and pMg. It can be shown that (dG ')T,P,pH,pMg 0, so that G ' is at a minimum when T, P, pH, and pMg are held constant. This is the fundamental justification for the use of G ' in biochemistry. Under the appropriate circumstances, the magnesium term can be left out or be replaced by a term in another metal ion. The reaction is generally an enzyme-catalyzed reaction, but these concepts apply to any reaction involving a weak acid or metal ion complex when the pH and concentration of free metal ion at equilibrium are specified.
A consequence of equation 46 is that the chemical potential i of each species in the system is replaced by the transformed chemical potential i ' given by
i ' = i - NH(i)(H+) - NMg(i)(Mg2+) . . . . . . . . (48)
where NH(i) is the number of hydrogen atoms in species i and NMg(i ) is the number of magnesium atoms in species i.
Although thermodynamic derivations are carried out using the chemical potential, in actual calculations, the chemical potential i of species i is replaced by the Gibbs energy of formation 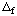 Gi and the transformed chemical potential i ' of species i is replaced by the transformed Gibbs energy of formation Gi ', where
Gi and the transformed chemical potential i ' of species i is replaced by the transformed Gibbs energy of formation Gi ', where
Gi = Gio + RT ln ([i ]/co) . . . . . . . . (49)
Gi ' = Gi 'o + RT ln ([i ]/co) . . . . . . . . (50)
for ideal solutions. The calculation of Gi 'o for a species is discussed in Section 5.4 and the calculation of Gi 'o for a reactant is discussed in Section 5.5.
Once the Gi 'o for the species (H2PO4-, HPO42-, MgHPO4) of Pi, for example, have been calculated, the next question is how can these values be combined to obtain the value of Gi 'o for Pi ? The equations for this calculation are given in the next section.
5.2 ISOMER GROUP THERMODYNAMICS
A problem that has to be faced in biochemical thermodynamics at specified pH and pMg is that a reactant may consist of various species in equilibrium at the specified pH and pMg. Fortunately, a group of isomers (or pseudoisomers) in equilibrium with each other have thermodynamic properties just like a species does, but we refer to the properties of a pseudoisomer group as transformed properties. The problem of calculating a standard transformed Gibbs energy of formation of a reactant like ATP also arises when a reactant exists in isomeric forms (or hydrated and unhydrated forms), even if it is not a weak acid and does not complex with metal ions, so first we discuss a simple isomerization. The thing that characterizes an isomer group in ideal solutions is that the distribution within the isomer group is a function of temperature only. For such solutions, the standard Gibbs energy of formation of an isomer group G o(iso) can be calculated from the standard Gibbs energies of formation Gi o of the various isomers using (ref. 23)
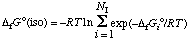 . . . . . . . . (51)
. . . . . . . . (51)
where NI is the number of isomers in the isomer group. The standard enthalpy of formation H o(iso) of the isomer group can be calculated using (ref. 24)
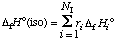 . . . . . . . . (52)
. . . . . . . . (52)
where ri is the equilibrium mole fraction of the i th species within the isomer group that is given by
ri = exp{[G o(iso) - Gi o]/RT} . . . . . . . . (53)
The standard entropy of formation of the isomer group S o(iso) is given by
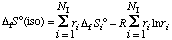 . . . . . . . . (54)
. . . . . . . . (54)
These equations can be used for pseudoisomer groups (for example, the species of ATP at specified pH and pMg) by using the transformed thermodynamic properties of the species.
For pseudoisomer groups, equations 51, 52, and 53 become
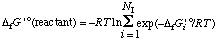 . . . . . . . . (55)
. . . . . . . . (55)
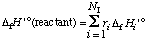 . . . . . . . . (56)
. . . . . . . . (56)
ri = exp{[G 'o(reactant) - Gi 'o]/RT} . . . . . . . . (57)
where i refers to a species at specified pH and specified free concentrations of metal ions that are bound.
5.3 ADJUSTMENT FOR IONIC STRENGTH
The ionic strength has a significant effect on the thermodynamic properties of ions, and the extended Debye-Huckel theory can be used to adjust the standard Gibbs energy of formation and the standard enthalpy of formation of ion i to the desired ionic strength (ref. 25-28). At 298.15 K these adjustments can be approximated by
Gi o(I) = Gi o(I = 0) - 2.91482zi2I 1/2/(1 + BI 1/2) . . . . . . . . (58)
Hi o(I) = Hi o(I = 0) + 1.4775zi2I 1/2/(1 + BI 1/2) . . . . . . . . (59)
where kJ mol-1 are used, zi is the charge on ion i, and B = 1.6 L1/2 mol-1/2. Since for H+, G o = 0 and H o = 0 at each temperature at I = 0, G o(H+, 298.15 K, I = 0.25 M) = -0.81 kJ mol-1 and H o(H+, 298.15 K, I = 0.25 M) = 0.41 kJ mol-1. For the purpose of these recommendations, pH = - log10([H+]/c o) and pMg = - log10([Mg2+]/c o), as discussed above in Section 4.1.
The adjustment of thermodynamic quantities from one solution composition to another using ionic strength effects alone is an approximation that works well at low ionic strengths (< 0.1 M) but it can fail at higher ionic strengths. Rigorous treatments require the use of interaction parameters (ref. 29) and a knowledge of the composition of the solution. While a substantial body of information on these parameters exists for aqueous inorganic solutions, there is very little of this type of data available for biochemical substances. Therefore, it is important that complete information on the compositions of the solutions used in equilibrium and calorimetric measurements be reported so that when values of these interaction parameters eventually become available, the results can be treated in a more rigorous manner. Specific ion effects are especially important when nucleic acids, proteins, and other polyelectrolytes are involved (refs. 30, 31).
5.4 ADJUSTMENT OF STANDARD THERMODYNAMIC PROPERTIES OF SPECIES TO THE DESIRED pH AND pMg
When pH and pMg are specified, the various species of ATP, for example, become pseudoisomers; that is their relative concentrations are a function of temperature only. The procedure for calculating the transformed chemical potential i ' of a species has been indicated in equation 48. For actual calculations the chemical potentials i of species are replaced with Gi (see equation 49), and the transformed chemical potentials i ' of species are replaced with Gi ' (see equation 50). Thus equation 48 for a species can be written
Gi 'o = Gio - NH(i )[G o(H+) + RTln([H+]/c o)] - NMg(i)[G o(Mg2+) + RT ln([Mg2+]/c o) . . . . . . . . (60)
where NH(i) is the number of hydrogen atoms in species i. The corresponding equation for the standard transformed enthalpy of formation of species i is
Hi 'o = Hio - NH(i )Ho(H+) - NMg(i )Ho(Mg2+) . . . . . . . . (61)
since the enthalpy of an ion in an ideal solution is independent of its concentration.
In adjusting standard Gibbs energies of formation to a specified pH, there is the question as to whether to count all of the hydrogens or only those involved in the reaction under consideration. However, the recommendation here is to adjust for all of the hydrogens in a species because all of them may be ultimately removed in biochemical reactions. This has been done in Tables II and III.
There is a simple way to look at the standard transformed Gibbs energy of formation Gi'o and the standard transformed enthalpy of formation Hi'o of species i, and that is that they are the changes in formation reactions of the species with H+ at the specified pH and Mg2+ at the specified pMg on the left-hand side of the formation reaction (ref. 32). For H2PO4-,
P(s) + 2O2(g) + 2H+(pH = 7) + 3e- = H2PO4- . . . . . . . . (62)
G 'o(H2PO4-) = G o(H2PO4-) - 2{G o(H+) + RT ln 10-pH} . . . . . . . . (63)
H 'o(H2PO4-) = H o(H2PO4-) - 2H o(H+) . . . . . . . . (64)
The quantities Go(H+) and H o(H+) are included because they are equal to zero only at zero ionic strength. The electrons required to balance the formation reaction are assigned G o(e-) = H o(e-) = 0. This calculation can be made with either the standard thermodynamic properties at I = 0 or at some specified ionic strength.
The calculation of G 'o and H 'o for HPO42- and MgHPO4 follow this same pattern, with Mg2+(pMg = 3) also on the left-hand side of the formation reaction of MgHPO4.
For a pseudoisomer group in which G o and Ho are not known for any species, zero values have to be assigned to one of the species, as described in Section 4.1.
5.5 CALCULATION OF THE STANDARD FORMATION PROPERTIES OF A PSEUDOISOMER GROUP AT SPECIFIED pH AND pMg
H2PO4-, HPO42- and MgHPO4 form a pseudoisomer group when pH and pMg are specified. Therefore, equations 55-57 can be used to calculate G 'o(Pi) and H 'o(Pi) for inorganic phosphate at the desired pH and pMg. These calculations have been made for inorganic phosphate and glucose 6-phosphate at pH = 7 and pMg = 3 by Alberty (ref. 4), and for adenosine, AMP, ADP, and ATP by Alberty and Goldberg (ref. 7) using the convention that G o = H o = 0 for neutral adenosine.
For less common and more complicated reactants, the acid dissociation constants and magnesium complex dissociation constants may not be known. The Gi 'o values of the reactants at pH = 7 and pMg = 3 can, however, be calculated if K ' has been measured at pH = 7 and pMg = 3 for a reaction in which Gi 'o is known for the other reactants. For example, this approach can be used to calculate Gi 'o for the reactants in glycolysis.
5.6 THE ACTUAL EXPERIMENT AND THOUGHT EXPERIMENTS
In the laboratory, a biochemical equilibrium experiment is actually carried out at specified T and P, and the pH is measured at equilibrium. Buffers are used to hold the pH constant, but there may be a change in the pH if the catalyzed reaction produces or consumes acid. pMg at equilibrium has to be calculated, and this can be done accurately only if the acid dissociation constants and magnesium complex dissociation constants are known for all of the reactants and buffer components (ref. 12, 33). In the absence of this information pMg can be calculated approximately if the buffer binds H+ and Mg2+, these dissociation constants are known, and the concentrations of the reactants are much smaller than the concentration of the buffer components that are primarily responsible for the binding of Mg2+. We can hope that some day there will be a pMg electrode as convenient as the pH electrode.
When we interpret the thermodynamics of a biochemical equilibrium experiment, we use an idealized thought experiment that is equivalent to the laboratory experiment. In the laboratory experiment, the buffer determines the approximate pH, but the pH will drift if H+ is produced or consumed. The pH should be measured at equilibrium because the composition and  G 'o and H 'o depend on this pH. Since the experimental results depend on the final pH, we can imagine that the experiment was carried out in a reaction vessel with a semipermeable membrane (permeable to H+ and an anion, and impermeable to other reactants) with a pH reservoir on the other side. If the binding of H+ by the products is greater than that of the reactants, H+ will diffuse in from the pH reservoir as the reaction proceeds. If the binding of H+ by the reactants is greater, H+ will diffuse out of the reaction vessel as the reaction proceeds. Thus hydrogen ion is not conserved in the reaction vessel in this idealized thought experiment. Similar statements can be made about Mg2+. In calorimetric experiments, corrections have to be made for the enthalpies of reaction due to the production of H+ and Mg2+ to obtain H 'o, as mentioned earlier.
G 'o and H 'o depend on this pH. Since the experimental results depend on the final pH, we can imagine that the experiment was carried out in a reaction vessel with a semipermeable membrane (permeable to H+ and an anion, and impermeable to other reactants) with a pH reservoir on the other side. If the binding of H+ by the products is greater than that of the reactants, H+ will diffuse in from the pH reservoir as the reaction proceeds. If the binding of H+ by the reactants is greater, H+ will diffuse out of the reaction vessel as the reaction proceeds. Thus hydrogen ion is not conserved in the reaction vessel in this idealized thought experiment. Similar statements can be made about Mg2+. In calorimetric experiments, corrections have to be made for the enthalpies of reaction due to the production of H+ and Mg2+ to obtain H 'o, as mentioned earlier.
The thermodynamic interpretation of the apparent equilibrium constant K ' uses G 'o and H 'o. These quantities correspond with another thought experiment in which the separated reactants, each at 1 M at the specified T, P, final pH, final pMg, and I react to form the separated products, each at 1 M at the specified T, P, final pH, final pMg, and I.
5.7 LINEAR ALGEBRA
It is generally understood that chemical equations conserve atoms and charge, but it is not generally known how the conservation equations for a chemical reaction system can be calculated from a set of chemical equations or how an independent set of chemical equations can be calculated from the conservation equations for the system. Nor is it well known that conservation equations in addition to atom and charge balances may arise from the mechanism of reaction. The quantitative treatment of conservation equations and chemical reactions requires the use of matrices and matrix operations (ref. 10, 23). When the equilibrium concentrations of species such as H+ and Mg2+ are specified, these species and electric charge are not conserved, and so a biochemical equation should not indicate that they are conserved. The current practice of using words like acetate and symbols like ATP and Pi is satisfactory provided that people understand the reason for using these words and symbols. It should be possible to distinguish between chemical equations and biochemical equations on sight, and this means that different symbols should be used for the reactants in these two types of equations.
A set of simple chemical equations has been discussed from the viewpoint of linear algebra (ref. 34). The hydrolysis of ATP to ADP and Pi at specified pH has also been discussed from the viewpoint of linear algebra which shows why the 4 chemical equations reduce down to a single biochemical equation (ref. 35).
The conservation matrix for a biochemical reaction is especially useful for the identification of the constraints in addition to element balances (ref. 36).
References for this section
4. Alberty, R. A. (1992) Biophys. Chem. 42, 117-131.
5. Alberty, R. A. (1992) Biophys. Chem. 43, 239-254.
7. Alberty, R. A., and Goldberg, R. N. (1992) Biochemistry 31, 10610-10615.
10. Alberty, R. A. (1992) J. Phys. Chem. 96, 9614-9621.
12. Teague, W. E., and Dobson, G. P. (1992) J. Biol. Chem. 267, 14084-14093.
20. Alberty, R. A., and Oppenheim, I. (1988) J. Chem. Phys. 89, 3689-3693.
21. Alberty, R. A., and Oppenheim, I. (1992) J. Chem. Phys. 96, 9050-9054.
22. Wyman, J., and Gill, S. J. (1990) Binding and Linkage, University Science Books, Mill Valley, CA.
23. Smith, W. R., and Missen, R. W. (1982) Chemical Reaction Equilibrium Analysis: Theory and Algorithms, Wiley-Interscience, New York.
24. Alberty, R. A. (1983) I & EC Fund. 22, 318-321.
25. Goldberg, R. N., and Tewari, Y. B. (1989) J. Phys. Chem. Ref. Data 18, 809-880.
26. Larson, J. W., Tewari, Y. B., and Goldberg, R. N. (1993) J. Chem. Thermodyn. 25, 73-90.
27. Goldberg, R. N., and Tewari, Y. B. (1991) Biophys. Chem. 40, 241-261.
28. Clarke, E. C. W., and Glew, D. N. (1980) J. Chem. Soc., Faraday Trans. 1 76, 1911-1916.
29. Pitzer, K. S. (1991) Ion Interaction Approach: Theory and Data Correlation, in Activity Coefficients in Electrolyte Solutions, 2nd Edition, K. S. Pitzer, editor, CRC Press, Boca Raton, Fla.
30. Record, M. T., Anderson, C. F., and Lohman, T. M. (1978) Q. Rev. Biophys. 11, 2.
31. Anderson, C. F., and Record, M. T. (1993) J. Phys. Chem. 97, 7116-7126.
32. Alberty, R. A. (1993) Pure Appl. Chem. 65, 883-888.
33. Guynn, R. W., and Veech, R. L. (1973) J. Biol. Chem. 248, 6966-6972.
34. Alberty, R. A. (1991) J. Chem. Educ. 68, 984.
35. Alberty, R. A. (1992) J. Chem. Educ. 69, 493.
36. Alberty, R. A. (1994) Biophys. Chem. 49, 251-261.
Continue with 6 Recommendations on Thermodynamic Tables.
Return to home page for Biochemical Thermodynamics.