Nomenclature Committee of the International Union of Biochemistry and Molecular Biology (NC-IUBMB)
Proposed Changes to the Enzyme List
The entries below are proposed additions and amendments to the Enzyme Nomenclature list. They were prepared for the NC-IUBMB by Kristian Axelsen, Sinéad Boyce, Richard Cammack, Ron Caspi, Minoru Kanehisa, Andrew McDonald, Gerry Moss, Dietmar Schomburg, Ida Schomburg and Keith Tipton. Comments and suggestions on these draft entries should be sent to Dr Andrew McDonald (Department of Biochemistry, Trinity College Dublin, Dublin 2, Ireland). The date on which an enzyme will be made official is appended after the EC number. To prevent confusion please do not quote new EC numbers until they are incorporated into the main list.
An asterisk before 'EC' indicates that this is an amendment to an existing enzyme rather than a new enzyme entry.
Contents
EC 1.1.1.298 3-hydroxypropionate dehydrogenase (NADP+) (4 December 2009)
EC 1.1.1.299 malate dehydrogenase [NAD(P)+] (4 December 2009)
EC 1.1.1.300 NADP-retinol dehydrogenase (4 December 2009)
EC 1.1.5.4 malate dehydrogenase (quinone) (4 December 2009)
EC 1.1.5.5 alcohol dehydrogenase (quinone) (4 December 2009)
EC 1.1.99.16 transferred now EC 1.1.5.4 (4 December 2009)
EC 1.2.1.75 malonyl CoA reductase (malonate semialdehyde-forming) (4 December 2009)
EC 1.2.1.76 succinate-semialdehyde dehydrogenase (acetylating) (4 December 2009)
EC 1.3.1.84 acrylyl-CoA reductase (NADPH) (4 December 2009)
EC 1.5.3.11 deleted, now EC 1.5.3.13, EC 1.5.3.14, EC 1.5.3.15, EC 1.5.3.16 and EC 1.5.3.17 (29 October 2009)
EC 1.5.3.13 N1-acetylpolyamine oxidase (29 October 2009)
EC 1.5.3.14 polyamine oxidase (propane-1,3-diamine-forming) (29 October 2009)
EC 1.5.3.15 N8-acetylspermidine oxidase (propane-1,3-diamine-forming) (29 October 2009)
EC 1.5.3.16 spermine oxidase (29 October 2009)
EC 1.5.3.17 non-specific polyamine oxidase (29 October 2009)
*EC 1.7.3.1 nitroalkane oxidase (4 December 2009)
EC 1.13.11.32 transferred now EC 1.13.12.16 (4 December 2009)
EC 1.13.12.16 nitronate monooxygenase (4 December 2009)
EC 1.14.14.7 tryptophan 7-halogenase (4 December 2009)
EC 2.1.1.163 demethylmenaquinone methyltransferase (4 December 2009)
EC 2.3.1.188 ω-hydroxypalmitate O-feruloyl transferase (23 November 2009)
EC 2.4.1.247 β-D-galactosyl-(1→4)-L-rhamnose phosphorylase (23 November 2009)
EC 2.4.1.248 cycloisomaltooligosaccharide glucanotransferase (4 December 2009)
EC 2.4.1.249 delphinidin 3',5'-O-glucosyltransferase (4 December 2009)
EC 2.4.2.41 xylogalacturonan β-1,3-xylosyltransferase (4 December 2009)
EC 2.5.1.8 transferred now EC 2.5.1.75 (4 December 2009)
EC 2.5.1.73 O-phospho-L-seryl-tRNA:Cys-tRNA synthase (4 December 2009)
EC 2.5.1.74 1,4-dihydroxy-2-naphthoate polyprenyltransferase (4 December 2009)
EC 2.5.1.75 tRNA dimethylallyltransferase (4 December 2009)
EC 2.5.1.76 cysteate synthase (4 December 2009)
*EC 2.7.1.119 hygromycin-B 7"-O-kinase (4 December 2009)
EC 2.7.1.163 hygromycin B 4-O-kinase (4 December 2009)
EC 2.7.1.164 O-phosphoseryl-tRNASec kinase (4 December 2009)
EC 2.9.1.2 O-phospho-L-seryl-tRNASec:L-selenocysteinyl-tRNA synthase (4 December 2009)
EC 3.1.26.13 retroviral ribonuclease H (4 December 2009)
EC 3.2.2.27 uracil-DNA glycosylase (4 December 2009)
EC 3.2.2.28 double-stranded uracil-DNA glycosylase (4 December 2009)
EC 3.2.2.29 thymine-DNA glycosylase (4 December 2009)
EC 3.4.17.23 angiotensin-converting enzyme 2 (4 December 2009)
EC 3.4.22.69 SARS coronavirus main proteinase (4 December 2009)
EC 3.4.22.70 sortase A (4 December 2009)
EC 3.4.22.71 sortase B (4 December 2009)
EC 3.4.23.50 human endogenous retrovirus K endopeptidase (4 December 2009)
EC 3.4.23.51 HycI peptidase (4 December 2009)
EC 3.4.24.87 ADAMTS13 endopeptidase (4 December 2009)
EC 3.4.25.2 HslU—HslV peptidase (4 December 2009)
EC 3.5.1.99 fatty acid amide hydrolase (23 November 2009)
EC 3.5.1.100 (R)-amidase (4 December 2009)
EC 3.5.1.101 L-proline amide hydrolase (4 December 2009)
EC 3.6.4.12 DNA helicase (4 December 2009)
EC 3.6.4.13 RNA helicase (4 December 2009)
EC 3.7.1.11 cyclohexane-1,2-dione hydrolase (4 December 2009)
EC 4.1.1.90 peptidyl-glutamate 4-carboxylase (4 December 2009)
EC 4.1.99.13 (6-4)DNA photolyase (23 November 2009)
EC 4.1.99.14 R-specific spore photoproduct lyase (4 December 2009)
EC 4.1.99.15 S-specific spore photoproduct lyase (4 December 2009)
EC 4.2.1.116 3-hydroxypropionyl-CoA dehydratase (4 December 2009)
EC 4.2.1.117 2-methylcitrate dehydratase (2-methyl-trans-aconitate forming) (4 December 2009)
EC 4.2.1.118 3-dehydroshikimate dehydratase (4 December 2009)
EC 4.2.1.119 enoyl-CoA hydratase 2 (4 December 2009)
EC 4.2.1.120 4-hydroxybutanoyl-CoA dehydratase (4 December 2009)
EC 4.2.3.45 phyllocladan-16α-ol synthase (12 November 2009)
EC 4.99.1.8 heme ligase (23 November 2009)
EC 6.1.1.27 O-phosphoserine—tRNA ligase (4 December 2009)
EC 6.2.1.36 3-hydroxypropionyl-CoA synthase (4 December 2009)
EC 6.3.1.13 L-cysteine:1D-myo-inositol 2-amino-2-deoxy-α-D-glucopyranoside ligase (23 November 2009)
EC 1.1.1.298
Accepted name: 3-hydroxypropionate dehydrogenase (NADP+)
Reaction: 3-hydroxypropionate + NADP+ = malonate semialdehyde + NADPH + H+
Systematic name: 3-hydroxypropionate:NADP+ oxidoreductase
Comments: Catalyses the reduction of malonate semialdehyde to 3-hydroxypropionate, a key step in the 3-hydroxypropionate and the 3-hydroxypropionate/4-hydroxybutyrate cycles, autotrophic CO2 fixation pathways found in some green non-sulfur phototrophic bacteria and archaea, respectively [1,2]. The enzyme from Chloroflexus aurantiacus is bifunctional, and also catalyses the upstream reaction in the pathway, EC 1.2.1.75 [3]. Different from EC 1.1.1.59 [3-hydroxypropionate dehydrogenase (NAD+)] by cofactor preference.
References:
1. Strauss, G. and Fuchs, G. Enzymes of a novel autotrophic CO2 fixation pathway in the phototrophic bacterium Chloroflexus aurantiacus, the 3-hydroxypropionate cycle. Eur. J. Biochem. 215 (1993) 633-643. [PMID: 8354269]
2. Berg, I.A., Kockelkorn, D., Buckel, W. and Fuchs, G. A 3-hydroxypropionate/4-hydroxybutyrate autotrophic carbon dioxide assimilation pathway in Archaea. Science 318 (2007) 1782-1786. [PMID: 18079405]
3. Hugler, M., Menendez, C., Schagger, H. and Fuchs, G. Malonyl-coenzyme A reductase from Chloroflexus aurantiacus, a key enzyme of the 3-hydroxypropionate cycle for autotrophic CO2 fixation. J. Bacteriol. 184 (2002) 2404-2410. [PMID: 11948153]
[EC 1.1.1.298 created 2009]
EC 1.1.1.299
Accepted name: malate dehydrogenase [NAD(P)+]
Reaction: (S)-malate + NAD(P)+ = oxaloacetate + NAD(P)H + H+
Other name(s): MdH II, NAD(P)+-dependent malate dehyrogenase
Systematic name: (S)-malate:NAD(P)+ oxidoreductase
Comments: This enzyme, which was characterized from the methanogenic archaeon Methanobacterium thermoautotrophicum, catalyses only the reduction of oxaloacetate, and can use NAD+ and NADP+ with similar specific activity [1]. Different from EC 1.1.1.37 [malate dehydrogenase (NAD+)], EC 1.1.1.82 [malate dehydrogenase (NADP+)] and EC 1.1.5.4 [malate dehydrogenase (quinone)].
References:
1. Thompson, H., Tersteegen, A., Thauer, R.K. and Hedderich, R. Two malate dehydrogenases in Methanobacterium thermoautotrophicum. Arch. Microbiol. 170 (1998) 38-42. [PMID: 9639601]
[EC 1.1.1.299 created 2009]
EC 1.1.1.300
Accepted name: NADP-retinol dehydrogenase
Reaction: retinol + NADP+ = retinal + NADPH + H+
Other name(s): all-trans retinal reductase; all-trans-retinol dehydrogenase; NADP(H)-dependent retinol dehydrogenase/reductase; RDH11; RDH12; RDH13; RDH14; retinol dehydrogenase 12; retinol dehydrogenase 14; retinol dehydrogenase [NADP+]; RalR1; PSDR1
Systematic name: retinol:NADP+ oxidoreductase
Comments: Greater catalytic efficiency in the reductive direction. This observation, and the enzyme's localization at the entrance to the mitochondrial matrix, suggest that it may function to protect mitochondria against oxidative stress associated with the highly reactive retinal produced from dietary β-carotene by EC 1.14.99.36 (β-carotene 15,15'-monooxygenase) [2]. Km-values for NADP+ and NADPH are at least 800-fold lower than those for NAD+ and NADH [1,4]. This enzyme differs from EC 1.1.1.105, retinol dehydrogenase, which prefers NAD+ and NADH. This enzyme is distinct from EC 1.1.1.71, alcohol dehydrogenase (NAD(P)+), which shows a broad specificity for aldehydes.
References:
1. Belyaeva, O.V., Korkina, O.V., Stetsenko, A.V., Kim, T., Nelson, P.S. and Kedishvili, N.Y. Biochemical properties of purified human retinol dehydrogenase 12 (RDH12): catalytic efficiency toward retinoids and C9 aldehydes and effects of cellular retinol-binding protein type I (CRBPI) and cellular retinaldehyde-binding protein (CRALBP) on the oxidation and reduction of retinoids. Biochemistry 44 (2005) 7035-7047. [PMID: 15865448]
2. Belyaeva, O.V., Korkina, O.V., Stetsenko, A.V. and Kedishvili, N.Y. Human retinol dehydrogenase 13 (RDH13) is a mitochondrial short-chain dehydrogenase/reductase with a retinaldehyde reductase activity. FEBS J. 275 (2008) 138-147. [PMID: 18039331]
3. Haeseleer, F., Huang, J., Lebioda, L., Saari, J.C. and Palczewski, K. Molecular characterization of a novel short-chain dehydrogenase/reductase that reduces all-trans-retinal. J. Biol. Chem. 273 (1998) 21790-21799. [PMID: 9705317]
4. Kedishvili, N.Y., Chumakova, O.V., Chetyrkin, S.V., Belyaeva, O.V., Lapshina, E.A., Lin, D.W., Matsumura, M. and Nelson, P.S. Evidence that the human gene for prostate short-chain dehydrogenase/reductase (PSDR1) encodes a novel retinal reductase (RalR1). J. Biol. Chem. 277 (2002) 28909-28915. [PMID: 12036956]
[EC 1.1.1.300 created 2009]
EC 1.1.5.4
Accepted name: malate dehydrogenase (quinone)
Reaction: (S)-malate + a quinone = oxaloacetate + reduced quinone
Other name(s): FAD-dependent malate-vitamin K reductase; malate-vitamin K reductase; (S)-malate:(acceptor) oxidoreductase; L-malate-quinone oxidoreductase; malate:quinone oxidoreductase; malate quinone oxidoreductase; MQO; malate:quinone reductase; malate dehydrogenase (acceptor); FAD-dependent malate dehydrogenase
Systematic name: (S)-malate:quinone oxidoreductase
Comments: A flavoprotein (FAD). Vitamin K and several other quinones can act as acceptors. Different from EC 1.1.1.37 [malate dehydrogenase (NAD+)], EC 1.1.1.82 [malate dehydrogenase (NADP+)] and EC 1.1.1.299 [malate dehydrogenase [NAD(P)+)].
References:
1. Imai, D. and Brodie, A.F. A phospholipid-requiring enzyme, malate-vitamin K reductase. J. Biol. Chem. 248 (1973) 7487-7494.
2. Imai, T. FAD-dependent malate dehydrogenase, a phospholipid-requiring enzyme from Mycobacterium sp. strain Takeo. Purification and some properties. Biochim. Biophys. Acta 523 (1978) 37-46. [PMID: 629992]
3. Reddy, T.L.P., Suryanarayana, P.M. and Venkitasubramanian, T.A. Variations in the pathways of malate oxidation and phosphorylation in different species of Mycobacteria. Biochim. Biophys. Acta 376 (1975) 210-218. [PMID: 234747]
4. Molenaar, D., van der Rest, M.E. and Petrovic, S. Biochemical and genetic characterization of the membrane-associated malate dehydrogenase (acceptor) from Corynebacterium glutamicum. Eur. J. Biochem. 254 (1998) 395-403. [PMID: 9660197]
5. Kather, B., Stingl, K., van der Rest, M.E., Altendorf, K. and Molenaar, D. Another unusual type of citric acid cycle enzyme in Helicobacter pylori: the malate:quinone oxidoreductase. J. Bacteriol. 182 (2000) 3204-3209. [PMID: 10809701]
[EC 1.1.5.4 created 1978 as EC 1.1.99.16, transferred 2009 to EC 1.1.5.4]
EC 1.1.5.5
Accepted name: alcohol dehydrogenase (quinone)
Reaction: ethanol + ubiquinone = acetaldehyde + ubiquinol
Other name(s): quinohaemoprotein alcohol dehydrogenase; PQQ ADH; PQQ alcohol dehydrogenase; PQQ dependent alcohol dehydrogenase; PQQ-alcohol dehydrogenase; PQQ-dependent ADH; PQQ-dependent alcohol dehydrogenase; pyrroloquinoline quinone-dependent alcohol dehydrogenase; pyrroloquinoline quinone-dependent quinoprotein methanol dehydrogenase; QH-ADH; quinohemoprotein alcohol dehydrogenases; quinoprotein ADH; quinoprotein alcohol dehydrogenase
Systematic name: alcohol:quinone oxidoreductase
Comments: In acetic acid bacteria this enzyme is involved in acetic acid production [3].
References:
1. Gomez-Manzo, S., Contreras-Zentella, M., Gonzalez-Valdez, A., Sosa-Torres, M., Arreguin-Espinoza, R. and Escamilla-Marvan, E. The PQQ-alcohol dehydrogenase of Gluconacetobacter diazotrophicus. Int. J. Food Microbiol. 125 (2008) 71-78. [PMID: 18321602]
2. Shinagawa, E., Toyama, H., Matsushita, K., Tuitemwong, P., Theeragool, G. and Adachi, O. A novel type of formaldehyde-oxidizing enzyme from the membrane of Acetobacter sp. SKU 14. Biosci. Biotechnol. Biochem. 70 (2006) 850-857. [PMID: 16636451]
3. Chinnawirotpisan, P., Theeragool, G., Limtong, S., Toyama, H., Adachi, O.O. and Matsushita, K. Quinoprotein alcohol dehydrogenase is involved in catabolic acetate production, while NAD-dependent alcohol dehydrogenase in ethanol assimilation in Acetobacter pasteurianus SKU1108. J. Biosci. Bioeng. 96 (2003) 564-571. [PMID: 16233574]
4. Frebortova, J., Matsushita, K., Arata, H. and Adachi, O. Intramolecular electron transport in quinoprotein alcohol dehydrogenase of Acetobacter methanolicus: a redox-titration stud. Biochim. Biophys. Acta 1363 (1998) 24-34. [PMID: 9526036]
5. Matsushita, K., Kobayashi, Y., Mizuguchi, M., Toyama, H., Adachi, O., Sakamoto, K. and Miyoshi, H. A tightly bound quinone functions in the ubiquinone reaction sites of quinoprotein alcohol dehydrogenase of an acetic acid bacterium, Gluconobacter suboxydans. Biosci. Biotechnol. Biochem. 72 (2008) 2723-2731. [PMID: 18838797]
[EC 1.1.5.5 created 2009]
[EC 1.1.99.16 Transferred entry: malate dehydrogenase (acceptor). As the acceptor is now known, the enzyme has been transferred to EC 1.1.5.4, malate dehydrogenase (quinone). (EC 1.1.99.16 created 1978, deleted 2009)]
EC 1.2.1.75
Accepted name: malonyl CoA reductase (malonate semialdehyde-forming)
Reaction: malonate semialdehyde + coenzyme A + NADP+ = malonyl-CoA + NADPH + H+
Other name(s): NADP-dependent malonyl CoA reductase; malonyl CoA reductase (NADP)
Systematic name: malonate semialdehyde:NADP+ oxidoreductase (malonate semialdehyde-forming)
Comments: Requires Mg2+. Catalyses the reduction of malonyl-CoA to malonate semialdehyde, a key step in the 3-hydroxypropionate and the 3-hydroxypropionate/4-hydroxybutyrate cycles, autotrophic CO2 fixation pathways found in some green non-sulfur phototrophic bacteria and some thermoacidophilic archaea, respectively [1,2]. The enzyme from Sulfolobus tokodaii has been purified, and found to contain one RNA molecule per two subunits [3]. The enzyme from Chloroflexus aurantiacus is bifunctional, and also catalyses the next reaction in the pathway, EC 1.1.1.298 [3-hydroxypropionate dehydrogenase (NADP+)] [4].
References:
1. Strauss, G. and Fuchs, G. Enzymes of a novel autotrophic CO2 fixation pathway in the phototrophic bacterium Chloroflexus aurantiacus, the 3-hydroxypropionate cycle. Eur. J. Biochem. 215 (1993) 633-643. [PMID: 8354269]
2. Berg, I.A., Kockelkorn, D., Buckel, W. and Fuchs, G. A 3-hydroxypropionate/4-hydroxybutyrate autotrophic carbon dioxide assimilation pathway in Archaea. Science 318 (2007) 1782-1786. [PMID: 18079405]
3. Alber, B., Olinger, M., Rieder, A., Kockelkorn, D., Jobst, B., Hugler, M. and Fuchs, G. Malonyl-coenzyme A reductase in the modified 3-hydroxypropionate cycle for autotrophic carbon fixation in archaeal Metallosphaera and Sulfolobus spp. J. Bacteriol. 188 (2006) 8551-8559. [PMID: 17041055]
4. Hugler, M., Menendez, C., Schagger, H. and Fuchs, G. Malonyl-coenzyme A reductase from Chloroflexus aurantiacus, a key enzyme of the 3-hydroxypropionate cycle for autotrophic CO2 fixation. J. Bacteriol. 184 (2002) 2404-2410. [PMID: 11948153]
[EC 1.2.1.75 created 2009]
EC 1.2.1.76
Accepted name: succinate-semialdehyde dehydrogenase (acetylating)
Reaction: succinate semialdehyde + coenzyme A + NADP+ = succinyl-CoA + NADPH + H+
Other name(s): succinyl-coA reductase; coenzyme-A-dependent succinate-semialdehyde dehydrogenase
Systematic name: succinate semialdehyde:NADP+ oxidoreductase (CoA-acetylating)
Comments: Catalyses the NADPH-dependent reduction of succinyl-CoA to succinate semialdehyde. The enzyme has been described in Clostridium kluyveri, where it participates in succinate fermentation [1]), and in Metallosphaera sedula, where it participates in the 3-hydroxypropionate/4-hydroxybutyrate cycle, an autotrophic CO2 fixation pathway found in some thermoacidophilic archaea [2,3].
References:
1. Sohling, B. and Gottschalk, G. Purification and characterization of a coenzyme-A-dependent succinate-semialdehyde dehydrogenase from Clostridium kluyveri. Eur. J. Biochem. 212 (1993) 121-127. [PMID: 8444151]
2. Alber, B., Olinger, M., Rieder, A., Kockelkorn, D., Jobst, B., Hugler, M. and Fuchs, G. Malonyl-coenzyme A reductase in the modified 3-hydroxypropionate cycle for autotrophic carbon fixation in archaeal Metallosphaera and Sulfolobus spp. J. Bacteriol. 188 (2006) 8551-8559. [PMID: 17041055]
3. Berg, I.A., Kockelkorn, D., Buckel, W. and Fuchs, G. A 3-hydroxypropionate/4-hydroxybutyrate autotrophic carbon dioxide assimilation pathway in Archaea. Science 318 (2007) 1782-1786. [PMID: 18079405]
[EC 1.2.1.76 created 2009]
EC 1.3.1.84
Accepted name: acrylyl-CoA reductase (NADPH)
Reaction: propanoyl-CoA + NADP+ = acrylyl-CoA + NADPH + H+
Glossary: acrylyl-Coa = acryloyl-CoA;
propionyl-CoA = propanoyl-CoA
Systematic name: propanoyl-CoA:NADP+ oxidoreductase
Comments: Catalyses a step in the 3-hydroxypropionate/4-hydroxybutyrate cycle, an autotrophic CO2 fixation pathway found in some thermoacidophilic archaea [1]. The reaction is catalysed in the opposite direction to that shown. The enzyme from Sulfolobus tokodaii does not act on either NADH or crotonyl-CoA [2]. Different from EC 1.3.1.8, which acts only on enoyl-CoA derivatives of carbon chain length 4 to 16. Contains Zn2+.
References:
1. Berg, I.A., Kockelkorn, D., Buckel, W. and Fuchs, G. A 3-hydroxypropionate/4-hydroxybutyrate autotrophic carbon dioxide assimilation pathway in Archaea. Science 318 (2007) 1782-1786. [PMID: 18079405]
2. Teufel, R., Kung, J.W., Kockelkorn, D., Alber, B.E. and Fuchs, G. 3-hydroxypropionyl-coenzyme A dehydratase and acryloyl-coenzyme A reductase, enzymes of the autotrophic 3-hydroxypropionate/4-hydroxybutyrate cycle in the Sulfolobales. J. Bacteriol. 191 (2009) 4572-4581. [PMID: 19429610]
[EC 1.3.1.84 created 2009]
[EC 1.5.3.11 Deleted entry: Now included with EC 1.5.3.13 N1-acetylpolyamine oxidase, EC 1.5.3.14 polyamine oxidase (propane-1,3-diamine-forming), EC 1.5.3.15 N8-acetylspermidine oxidase (propane-1,3-diamine-forming), EC 1.5.3.16 spermine oxidase and EC 1.5.3.17 non-specific polyamine oxidase (EC 1.5.3.11 created 1992, deleted 2009)]
EC 1.5.3.13
Accepted name: N1-acetylpolyamine oxidase
Reaction: (1) N1-acetylspermidine + O2 + H2O = putrescine + 3-acetamidopropanal + H2O2
(2) N1-acetylspermine + O2 + H2O = spermidine + 3-acetamidopropanal + H2O2
Other name(s): N1-acetylpolyamine oxidase; hPAO-1; PAO (ambiguous); mPAO; hPAO; polyamine oxidase (ambiguous)
Systematic name: N1-acetylpolyamine:oxygen oxidoreductase (3-acetamidopropanal-forming)
Comments: The enzyme also catalyses the reaction: N1,N12-diacetylspermine + O2 + H2O = N1-acetylspermidine + 3-acetamamidopropanal + H2O2 [1]. No or very weak activity with spermine, or spermidine in absence of aldehydes. In presence of aldehydes the enzyme catalyses the reactions: 1. spermine + O2 + H2O = spermidine + 3-aminopropanal + H2O2, and with weak efficiency 2. spermidine + O2 + H2O = putrescine + 3-aminopropanal + H2O2 [2]. A flavoprotein (FAD). This enzyme, encoded by the PAOX gene, is found in mammalian peroxisomes and oxidizes N1-acetylated polyamines at the exo (three-carbon) side of the secondary amine, forming 3-acetamamidopropanal. Since the products of the reactions are deacetylated polyamines, this process is known as polyamine back-conversion. Differs in specificity from EC 1.5.3.14 [polyamine oxidase (propane-1,3-diamine-forming)], EC 1.5.3.15 [N8-acetylspermidine oxidase (propane-1,3-diamine-forming)], EC 1.5.3.16 (spermine oxidase) and EC 1.5.3.17 (non-specific polyamine oxidase).
References:
1. Vujcic, S., Liang, P., Diegelman, P., Kramer, D.L. and Porter, C.W. Genomic identification and biochemical characterization of the mammalian polyamine oxidase involved in polyamine back-conversion. Biochem. J. 370 (2003) 19-28. [PMID: 12477380]
2. Jarvinen, A., Grigorenko, N., Khomutov, A.R., Hyvonen, M.T., Uimari, A., Vepsalainen, J., Sinervirta, R., Keinanen, T.A., Vujcic, S., Alhonen, L., Porter, C.W. and Janne, J. Metabolic stability of α-methylated polyamine derivatives and their use as substitutes for the natural polyamines. J. Biol. Chem. 280 (2005) 6595-6601. [PMID: 15611107]
3. Wang, Y., Hacker, A., Murray-Stewart, T., Frydman, B., Valasinas, A., Fraser, A.V., Woster, P.M. and Casero, R.A., Jr. Properties of recombinant human N1-acetylpolyamine oxidase (hPAO): potential role in determining drug sensitivity. Cancer Chemother Pharmacol 56 (2005) 83-90. [PMID: 15791459]
4. Wu, T., Yankovskaya, V. and McIntire, W.S. Cloning, sequencing, and heterologous expression of the murine peroxisomal flavoprotein, N1-acetylated polyamine oxidase. J. Biol. Chem. 278 (2003) 20514-20525. [PMID: 12660232]
[EC 1.5.3.13 created 2009]
EC 1.5.3.14
Accepted name: polyamine oxidase (propane-1,3-diamine-forming)
Reaction: spermidine + O2 + H2O = propane-1,3-diamine + 4-aminobutanal + H2O2
Other name(s): MPAO; maize PAO
Systematic name: spermidine:oxygen oxidoreductase (propane-1,3-diamine-forming)
Comments: As the products of the reaction cannot be converted directly to other polyamines, this class of polyamine oxidases is considered to be involved in the terminal catabolism of polyamines [1]. This enzyme less efficiently catalyses the oxidation of N1-acetylspermine and spermine. A flavoprotein (FAD). Differs in specificity from EC 1.5.3.13 (N1-acetylpolyamine oxidase), EC 1.5.3.15 [N8-acetylspermidine oxidase (propane-1,3-diamine-forming)], EC 1.5.3.16 (spermine oxidase) and EC 1.5.3.17 (non-specific polyamine oxidase).
References:
1. Tavladoraki, P., Schinina, M.E., Cecconi, F., Di Agostino, S., Manera, F., Rea, G., Mariottini, P., Federico, R. and Angelini, R. Maize polyamine oxidase: primary structure from protein and cDNA sequencing. FEBS Lett. 426 (1998) 62-66. [PMID: 9598979]
2. Federico, R., Ercolini, L., Laurenzi, M., Angelini, R. Oxidation of acetylpolyamines by maize polyamine oxidase. Phytochemistry 43 (1996) 339-341.
[EC 1.5.3.14 created 2009]
EC 1.5.3.15
Accepted name: N8-acetylspermidine oxidase (propane-1,3-diamine-forming)
Reaction: N8-acetylspermidine + O2 + H2O = propane-1,3-diamine + 4-acetamidobutanal + H2O2
Systematic name: N8-acetylspermidine:oxygen oxidoreductase (propane-1,3-diamine-forming)
Comments: Also active with N1-acetylspermine, weak activity with N1,N12-diacetylspermine. No activity with diaminopropane, putrescine, cadaverine, diaminohexane, norspermidine, spermine and spermidine. Absence of monoamine oxidase (EC 1.4.3.4) activity. Differs in specificity from EC 1.5.3.13 (N1-acetylpolyamine oxidase), EC 1.5.3.14 [polyamine oxidase (propane-1,3-diamine-forming)], EC 1.5.3.16 (spermine oxidase) and EC 1.5.3.17 (non-specific polyamine oxidase).
References:
1. Shukla, O.P., Muller, S. and Walter, R.D. Polyamine oxidase from Acanthamoeba culbertsoni specific for N8-acetylspermidine. Mol. Biochem. Parasitol. 51 (1992) 91-98. [PMID: 1565141]
[EC 1.5.3.15 created 2009]
EC 1.5.3.16
Accepted name: spermine oxidase
Reaction: spermine + O2 + H2O = spermidine + 3-aminopropanal + H2O2
Other name(s): PAOh1/SMO; PAOh1 (ambiguous); AtPAO1; AtPAO4; SMO; mSMO; SMO(PAOh1); SMO/PAOh1; SMO5; mSMOmu
Systematic name: spermidine:oxygen oxidoreductase (spermidine-forming)
Comments: The enzyme from Arabidopsis thaliana (AtPAO1) oxidizes norspermine to norspermidine with high efficiency [3]. The mammalian enzyme, encoded by the SMOX gene, is a cytosolic enzyme that catalyses the oxidation of spermine at the exo (three-carbon) side of the tertiary amine. No activity with spermidine. Weak activity with N1-acetylspermine. A flavoprotein (FAD). Differs in specificity from EC 1.5.3.13 (N1-acetylpolyamine oxidase), EC 1.5.3.14 [polyamine oxidase (propane-1,3-diamine-forming)], EC 1.5.3.15 [N8-acetylspermidine oxidase (propane-1,3-diamine-forming)] and EC 1.5.3.17 (non-specific polyamine oxidase).
References:
1. Murray-Stewart, T., Wang, Y., Goodwin, A., Hacker, A., Meeker, A. and Casero, R.A., Jr. Nuclear localization of human spermine oxidase isoforms - possible implications in drug response and disease etiology. FEBS J. 275 (2008) 2795-2806. [PMID: 18422650]
2. Cervelli, M., Polticelli, F., Federico, R. and Mariottini, P. Heterologous expression and characterization of mouse spermine oxidase. J. Biol. Chem. 278 (2003) 5271-5276. [PMID: 12458219]
3. Tavladoraki, P., Rossi, M.N., Saccuti, G., Perez-Amador, M.A., Polticelli, F., Angelini, R. and Federico, R. Heterologous expression and biochemical characterization of a polyamine oxidase from Arabidopsis involved in polyamine back conversion. Plant Physiol. 141 (2006) 1519-1532. [PMID: 16778015]
4. Wang, Y., Murray-Stewart, T., Devereux, W., Hacker, A., Frydman, B., Woster, P.M. and Casero, R.A., Jr. Properties of purified recombinant human polyamine oxidase, PAOh1/SMO. Biochem. Biophys. Res. Commun. 304 (2003) 605-611. [PMID: 12727196]
[EC 1.5.3.16 created 2009]
EC 1.5.3.17
Accepted name: non-specific polyamine oxidase
Reaction: (1) spermine + O2 + H2O = spermidine + 3-aminopropanal + H2O2
(2) spermidine + O2 + H2O = putrescine + 3-aminopropanal + H2O2
(3) N1-acetylspermine + O2 + H2O = spermidine + 3-acetamidopropanal + H2O2
(4) N1-acetylspermidine + O2 + H2O = putrescine + 3-acetamidopropanal + H2O2
Other name(s): polyamine oxidase (ambiguous); Fms1; AtPAO3
Systematic name: polyamine:oxygen oxidoreductase (3-aminopropanal or 3-acetamidopropanal-forming)
Comments: A flavoprotein (FAD). The non-specific polyamine oxidases may differ from each other considerably. The enzyme from Saccharomyces cerevisiae shows a rather broad specificity and also oxidizes N8-acetylspermidine [3]. The enzyme from Ascaris suum shows high activity with spermine and spermidine, but also oxidizes norspermine [2]. The enzyme from Arabidopsis thaliana shows high activity with spermidine, but also oxidizes other polyamines [1].
The specific polyamine oxidases are classified as EC 1.5.3.13 (N1-acetylpolyamine oxidase), EC 1.5.3.14 [polyamine oxidase (propane-1,3-diamine-forming)], EC 1.5.3.15 (N8-acetylspermidine oxidase (propane-1,3-diamine-forming)) and EC 1.5.3.16 (spermine oxidase).
References:
1. Moschou, P.N., Sanmartin, M., Andriopoulou, A.H., Rojo, E., Sanchez-Serrano, J.J. and Roubelakis-Angelakis, K.A. Bridging the gap between plant and mammalian polyamine catabolism: a novel peroxisomal polyamine oxidase responsible for a full back-conversion pathway in Arabidopsis. Plant Physiol. 147 (2008) 1845-1857. [PMID: 18583528]
2. Muller, S. and Walter, R.D. Purification and characterization of polyamine oxidase from Ascaris suum. Biochem. J. 283 (1992) 75-80. [PMID: 1567380]
3. Landry, J. and Sternglanz, R. Yeast Fms1 is a FAD-utilizing polyamine oxidase. Biochem. Biophys. Res. Commun. 303 (2003) 771-776. [PMID: 12670477]
[EC 1.5.3.17 created 2009]
*EC 1.7.3.1
Accepted name: nitroalkane oxidase
Reaction: a nitroalkane + H2O + O2 = an aldehyde or ketone + nitrite + H2O2
Other name(s): nitroethane oxidase; NAO; nitroethane:oxygen oxidoreductase
Systematic name: nitroalkane:oxygen oxidoreductase
Comments: Has an absolute requirement for FAD [4]. While nitroethane may be the physiological substrate [2], the enzyme also acts on several other nitroalkanes, including 1-nitropropane, 2-nitropropane, 1-nitrobutane, 1-nitropentane, 1-nitrohexane, nitrocyclohexane and some nitroalkanols [4]. Differs from EC 1.13.11.16, nitronate monooxygenase, in that the preferred substrates are neutral nitroalkanes rather than anionic nitronates [4].
Links to other databases:
BRENDA,
EXPASY,
KEGG, PDB,
CAS registry number: 9029-36-1, 65802-82-6
References:
1. Little, H.N. Oxidation of nitroethane by extracts from Neurospora. J. Biol. Chem. 193 (1951) 347-358. [PMID: 14907722]
2. Kido, T., Hashizume, K. and Soda, K. Purification and properties of nitroalkane oxidase from Fusarium oxysporum. J. Bacteriol. 133 (1978) 53-58. [PMID: 22538]
3. Daubner, S.C., Gadda, G., Valley, M.P. and Fitzpatrick, P.F. Cloning of nitroalkane oxidase from Fusarium oxysporum identifies a new
member of the acyl-CoA dehydrogenase superfamily. Proc. Natl. Acad. Sci. USA 99 (2002) 2702-2707. [PMID: 11867731]
4. Fitzpatrick, P.F., Orville, A.M., Nagpal, A. and Valley, M.P. Nitroalkane oxidase, a carbanion-forming flavoprotein homologous to
acyl-CoA dehydrogenase. Arch. Biochem. Biophys. 433 (2005) 157-165. [PMID: 15581574]
5. Valley, M.P., Tichy, S.E. and Fitzpatrick, P.F. Establishing the kinetic competency of the cationic imine intermediate in
nitroalkane oxidase. J. Am. Chem. Soc. 127 (2005) 2062-2066. [PMID: 15713081]
[EC 1.7.3.1 created 1961, modified 2006, modified 2009]
[EC 1.13.11.32 Transferred entry: 2-nitropropane dioxygenase. Now EC 1.13.12.16, nitronate monooxygenase. (EC 1.13.11.32 created 1984, modified 2006, deleted 2009)]
EC 1.13.12.16
Accepted name: nitronate monooxygenase
Reaction: ethylnitronate + O2 + FMNH2 = acetaldehyde + nitrite + FMN + H2O
Other name(s): NMO; 2-nitropropane dioxygenase (incorrect)
Systematic name: nitronate:oxygen 2-oxidoreductase (nitrite-forming)
Comments: Previously classified as 2-nitropropane dioxygenase (EC 1.13.11.32) [1,2], but it is now recognized that this was the result of the slow ionization of nitroalkanes to their nitronate (anionic) forms [3]. The enzymes from the fungus Neurospora crassa and the yeast Williopsis saturnus var. mrakii (formerly classified as Hansenula mrakii) contain non-covalently bound FMN as the cofactor. Active towards linear alkyl nitronates of lengths between 2 and 6 carbon atoms and, with lower activity, towards propyl-2-nitronate. The enzyme from N. crassa can also utilize neutral nitroalkanes, but with lower activity. One atom of oxygen is incorporated into the carbonyl group of the aldehyde product. The reaction appears to involve the formation of an enzyme-bound nitronate radical and an α-peroxynitroethane species, which then decomposes, either in the active site of the enzyme or after release, to acetaldehyde and nitrite. A flavoprotein (FMN).
References:
1. Francis, K., Russell, B. and Gadda, G. Involvement of a flavosemiquinone in the enzymatic oxidation of
nitroalkanes catalyzed by 2-nitropropane dioxygenase. J. Biol. Chem. 280 (2005) 5195-5204. [PMID: 15582992]
2. Ha, J.Y., Min, J.Y., Lee, S.K., Kim, H.S., Kim do, J., Kim, K.H., Lee, H.H., Kim, H.K., Yoon, H.J. and Suh, S.W. Crystal structure of 2-nitropropane dioxygenase complexed with FMN and substrate. Identification of the catalytic base. J. Biol. Chem. 281 (2006) 18660-18667. [PMID: 16682407]
3. Gadda, G. and Francis, K. Nitronate monooxygenase, a model for anionic flavin semiquinone intermediates in oxidative catalysis. Arch. Biochem. Biophys. (2009) . [PMID: 19577534]
4. Francis, K. and Gadda, G. Kinetic evidence for an anion binding pocket in the active site of nitronate monooxygenase. Bioorg. Chem. 37 (2009) 167-172. [PMID: 19683782]
[EC 1.13.12.16 created 1984 as EC 1.13.11.32, modified 2006, transferred 2009 to EC 1.13.12.16]
EC 1.14.14.7
Accepted name: tryptophan 7-halogenase
Reaction: tryptophan + FADH2 + Cl- + O2 + H+ = 7-chloro-L-tryptophan + FAD + 2 H2O
Other name(s): PrnA; RebH
Systematic name: L-tryptophan:FADH2 oxidoreductase (7-halogenating)
Comments: In Lechevalieria aerocolonigenes the enzyme catalyses the initial step in the biosynthesis of rebeccamycin [2].
References:
1. Dong, C., Kotzsch, A., Dorward, M., van Pee, K.H. and Naismith, J.H. Crystallization and X-ray diffraction of a halogenating enzyme, tryptophan 7-halogenase, from Pseudomonas fluorescens. Acta Crystallogr. D Biol. Crystallogr. 60 (2004) 1438-1440. [PMID: 15272170]
2. Yeh, E., Garneau, S. and Walsh, C.T. Robust in vitro activity of RebF and RebH, a two-component reductase/halogenase, generating 7-chlorotryptophan during rebeccamycin biosynthesis. Proc. Natl. Acad. Sci. USA 102 (2005) 3960-3965. [PMID: 15743914]
3. Bitto, E., Huang, Y., Bingman, C.A., Singh, S., Thorson, J.S. and Phillips Jr., G.N. The structure of flavin-dependent tryptophan 7-halogenase RebH. Proteins Struct. Funct. Genet. 70 (2008) 289-293.
[EC 1.14.14.7 created 2009]
EC 2.1.1.163
Accepted name: demethylmenaquinone methyltransferase
Reaction: a demethylmenaquinol + S-adenosyl-L-methionine = a menaquinol + S-adenosyl-L-homocysteine
Other name(s): S-adenosyl-L-methione—DMK methyltransferase; demethylmenaquinone C-methylase; 2-heptaprenyl-1,4-naphthoquinone methyltransferase; 2-demethylmenaquinone methyltransferase; S-adenosyl-L-methione:2-demethylmenaquinone methyltransferase
Systematic name: S-adenosyl-L-methione:demethylmenaquinone methyltransferase
Comments: The enzyme catalyses the last step in menaquinone biosynthesis. It is able to accept substrates with varying polyprenyl side chain length (the chain length is determined by polyprenyl diphosphate synthase)[1]. The enzyme from Escherichia coli also catalyses the conversion of 2-methoxy-6-octaprenyl-1,4-benzoquinone to 5-methoxy-2-methyl-3-octaprenyl-1,4-benzoquinone during the biosynthesis of ubiquinone [4]. The enzyme probably acts on menaquinol rather than menaquinone.
References:
1. Koike-Takeshita, A., Koyama, T. and Ogura, K. Identification of a novel gene cluster participating in menaquinone (vitamin K2) biosynthesis. Cloning and sequence determination of the 2-heptaprenyl-1,4-naphthoquinone methyltransferase gene of Bacillus stearothermophilus. J. Biol. Chem. 272 (1997) 12380-12383. [PMID: 9139683]
2. Wissenbach, U., Ternes, D. and Unden, G. An Escherichia coli mutant containing only demethylmenaquinone, but no menaquinone: effects on fumarate, dimethylsulfoxide, trimethylamine N-oxide and nitrate respiration. Arch. Microbiol. 158 (1992) 68-73. [PMID: 1444716]
3. Catala, F., Azerad, R. and Lederer, E. Sur les propriétés de la desméthylménaquinone C-méthylase de Mycobacterium phlei. Int. Z. Vitaminforsch. 40 (1970) 363-373. [PMID: 5450997]
4. Lee, P.T., Hsu, A.Y., Ha, H.T. and Clarke, C.F. A C-methyltransferase involved in both ubiquinone and menaquinone biosynthesis: isolation and identification of the Escherichia coli ubiE gene. J. Bacteriol. 179 (1997) 1748-1754. [PMID: 9045837]
[EC 2.1.1.163 created 2009]
EC 2.3.1.188
Accepted name: ω-hydroxypalmitate O-feruloyl transferase
Reaction: feruloyl-CoA + 16-hydroxypalmitate = CoA + 16-feruloyloxypalmitate
Glossary: 16-feruloyloxypalmitate = 16-{[3-(4-hydroxy-3-methoxyphenyl)prop-2-enoyl]oxy}hexadecanoate
Other name(s): hydroxycinnamoyl-CoA ω-hydroxypalmitic acid O-hydroxycinnamoyltransferase; HHT
Systematic name: feruloyl-CoA:16-hydroxypalmitate feruloyltransferase
Comments: p-Coumaroyl-CoA and sinapoyl-CoA also act as substrates. The enzyme is widely distributed in roots of higher plants.
References:
1. Lotfy, S., Negrel, J. and Javelle, F. Formation of feruloyloxypalmitic acid by an enzyme from wound-healing potato tuber discs. Phytochemistry 35 (1994) 1419-1424.
2. Lotfy, S. Javelle, F. and Negrel, J. Distribution of hydroxycinnamoyl-CoA ω-hydroxypalmitic acid O-hydroxycinnamoyltransferase in higher plants. Phytochemistry 40 (1995) 389-391.
3. Lotfy, S. Javelle, F. and Negrel, J. Purification and characterization of hydroxycinnamoyl-CoA ω-hydroxypalmitic acid O-hydroxycinnamoyltransferase from tobacco (Nicotiana tabacum L.) cell-suspension cultures. Planta 199 (1996) 475-480.
[EC 2.3.1.188 created 2009]
EC 2.4.1.247
Accepted name: β-D-galactosyl-(1→4)-L-rhamnose phosphorylase
Reaction: β-D-galactosyl-(1→4)-L-rhamnose + phosphate = L-rhamnose + α-D-galactose 1-phosphate
Other name(s): D-galactosyl-β1→4-L-rhamnose phosphorylase; GalRhaP
Systematic name: β-D-galactosyl-(1→4)-L-rhamnose:phosphate 1-α-D-galactosyltransferase
Comments: The enzyme from Clostridium phytofermentans is also active towards towards β-D-galactosyl derivatives of L-mannose, L-lyxose, D-glucose, 2-deoxy-D-glucose, and D-galactose in this order. Differs from 1,3-β-galactosyl-N-acetylhexosamine phosphorylase (EC 2.4.1.211) in being active towards L-rhamnose and inactive towards N-acetyl hexosamine derivatives.
References:
1. Nakajima, M., Nishimoto, M. and Kitaoka, M. Characterization of three β-galactoside phosphorylases from Clostridium phytofermentans: discovery of D-galactosyl-β1→4-L-rhamnose phosphorylase. J. Biol. Chem. 284 (2009) 19220-19227. [PMID: 19491100]
[EC 2.4.1.247 created 2009]
EC 2.4.1.248
Accepted name: cycloisomaltooligosaccharide glucanotransferase
Reaction: cyclizes part of a (1→6)-α-D-glucan chain by formation of a (1→6)-α-D-glucosidic bond
Systematic name: (1→6)-α-D-glucan:(1→6)-α-D-glucan 6-α-D-[1→6α-D-glucano]-transferase (cyclizing)
Comments: Specific for (1→6)-α-D-glucans (dextrans) and, unlike cyclomaltodextrin glucanotransferase (EC 2.4.1.19), without activity towards (1→4)-α-D-glucans, such as amylose. It also has no activity on oligosaccharides, such as amylopectin and pullulan, containing (1→6)-α-D-glucosidic linkages at branch points. The enzyme from Bacillus circulans T-3040 has been shown to form cycloisomalto-oligosaccharides of three sizes (7, 8 and 9 glucose units). It will also catalyse the disproportionation of two isomalto-oligosaccharides molecules to yield a series of isomalto-oligosachharides and the addition of D-glucose to cycloisomalto-oligosaccharides with ring opening to form isomalto-oligosaccharides.
References:
1. Oguma T, Horiuchi, T, and Kobayashi M. Novel Cyclic dextrins, Cycloisomaltooligosaccharides, from Bacillus sp. T-3040 culture. Biosci. Biotech. Biochem. 57 (1993) 1225-1227.
2. Oguma, T., Tobe, K. and Kobayashi, M. Purification and properties of a novel enzyme from Bacillus spp. T-3040, which catalyzes the conversion of dextran to cyclic isomaltooligosaccharides. FEBS Lett. 345 (1994) 135-138. [PMID: 7515357]
3. Yamamoto, T., Terasawa, K., Kim, Y.M., Kimura, A., Kitamura, Y., Kobayashi, M. and Funane, K. Identification of catalytic amino acids of cyclodextran glucanotransferase from Bacillus circulans T-3040. Biosci. Biotechnol. Biochem. 70 (2006) 1947-1953. [PMID: 16926507]
[EC 2.4.1.248 created 2009]
EC 2.4.1.249
Accepted name: delphinidin 3',5'-O-glucosyltransferase
Reaction: (1a) UDP-glucose + delphinidin 3-O-(6"-O-malonyl)-β-D-glucoside = UDP + delphinidin 3-O-(6"-O-malonyl)-β-D-glucoside-3'-O-β-D-glucoside
(1b) UDP-glucose + delphinidin 3-O-(6"-O-malonyl)-β-D-glucoside-3'-O-β-D-glucoside = UDP + delphinidin 3-O-(6"-O-malonyl)-β-D-glucoside-3',5'-di-O-β-D-glucoside
Glossary: delphinidin 3-O-(6"-O-malonyl)-β-D-glucoside-3',5'-di-O-β-D-glucoside = ternatin C5.
Other name(s): UDP-glucose:anthocyanin 3',5'-O-glucosyltransferase; UA3'5’GZ
Systematic name: UDP-glucose:delphinidin 3-O-(6"-O-malonyl)-β-D-glucoside 3'-O-glucosyltransferase
Comments: Ternatins are a group of polyacetylated delphinidin glucosides that confer
blue color to the petals of butterfly pea (Clitoria ternatea).
This enzyme catalyses two reactions in the biosynthesis of ternatin C5: the conversion of delphinidin 3-O-(6"-O-malonyl)-β-D-glucoside to delphinidin 3-O-(6"-O-malonyl)-β-D-glucoside-3'-O-β-D-glucoside, followed by the conversion of the later to ternatin C5, by transferring two glucosyl groups in a stepwise manner [1].
References:
1. Kogawa, K., Kato, N., Kazuma, K., Noda, N. and Suzuki, M. Purification and characterization of UDP-glucose: anthocyanin 3',5'-O-glucosyltransferase from Clitoria ternatea. Planta 226 (2007) 1501-1509. [PMID: 17668234]
[EC 2.4.1.249 created 2009]
EC 2.4.2.41
Accepted name: xylogalacturonan β-1,3-xylosyltransferase
Reaction: Transfers a xylosyl residue from UDP-D-xylose to a D-galactose residue in xylogalacturonan, forming a β-1,3-D-xylosyl-D-galactose linkage.
Other name(s): xylogalacturonan xylosyltransferase; XGA xylosyltransferase
Systematic name: UDP-D-xylose:xylogalacturonan β-1,3-xylosyltransferase
Comments: Involved in plant cell wall synthesis. The enzyme from Arabidopsis thaliana could transfer D-xylose from UDP-D-xylose onto oligogalacturonide acceptors. The enzyme did not show significant activity with UDP-glucose, UDP-galactose, or UDP-N-acetyl-D-glucosamine as sugar donors.
References:
1. Jensen, J.K., Sorensen, S.O., Harholt, J., Geshi, N., Sakuragi, Y., Moller, I., Zandleven, J., Bernal, A.J., Jensen, N.B., Sorensen, C., Pauly, M., Beldman, G., Willats, W.G. and Scheller, H.V. Identification of a xylogalacturonan xylosyltransferase involved in pectin biosynthesis in Arabidopsis. Plant Cell 20 (2008) 1289-1302. [PMID: 18460606]
[EC 2.4.2.41 created 2009]
[EC 2.5.1.8 Transferred entry: tRNA isopentenyltransferase. As it is now known that the substrate is dimethyallyl diphosphate, the enzyme has been transferred to EC 2.5.1.75, tRNA dimethylallyltransferase. (EC 2.5.1.8 created 1972, deleted 2009)]
EC 2.5.1.73
Accepted name: O-phospho-L-seryl-tRNA:Cys-tRNA synthase
Reaction: O-phospho-L-seryl-tRNACys + sulfide = L-cysteinyl-tRNACys + phosphate
Other name(s): SepCysS; Sep-tRNA:Cys-tRNA synthase
Systematic name: O-phospho-L-seryl-tRNACys:hydrogen sulfide 2-aminopropanoate transferase
Comments: In organisms like Archaeoglobus fulgidus lacking EC 6.1.1.16 (cysteine—tRNA ligase) for the direct Cys-tRNACys formation, Cys-tRNACys is produced by an indirect pathway, in which EC 6.1.1.27 (O-phosphoseryl-tRNA ligase) ligates O-phosphoserine to tRNACys, and EC 2.5.1.73 converts the produced O-phospho-L-seryl-tRNACys to Cys-tRNACys. The SepRS/SepCysS pathway is the sole route for cysteine biosynthesis in the organism [1]. Methanosarcina mazei can use both pathways, the direct route using EC 6.1.1.16 (cysteine—tRNA ligase) and the indirect pathway with EC 6.1.1.27 (O-phosphoseryl-tRNA ligase) and EC 2.5.1.73 [2].
References:
1. Fukunaga, R. and Yokoyama, S. Structural insights into the second step of RNA-dependent cysteine biosynthesis in archaea: crystal structure of Sep-tRNA:Cys-tRNA synthase from Archaeoglobus fulgidus. J. Mol. Biol. 370 (2007) 128-141. [PMID: 17512006]
2. Hauenstein, S.I. and Perona, J.J. Redundant synthesis of cysteinyl-tRNACys in Methanosarcina mazei. J. Biol. Chem. 283 (2008) 22007-22017. [PMID: 18559341]
3. Yuan, J., Palioura, S., Salazar, J.C., Su, D., O'Donoghue, P., Hohn, M.J., Cardoso, A.M., Whitman, W.B. and Soll, D. RNA-dependent conversion of phosphoserine forms selenocysteine in eukaryotes and archaea. Proc. Natl. Acad. Sci. USA 103 (2006) 18923-18927. [PMID: 17142313]
[EC 2.5.1.73 created 2009]
EC 2.5.1.74
Accepted name: 1,4-dihydroxy-2-naphthoate polyprenyltransferase
Reaction: an all-trans-polyprenyl diphosphate + 1,4-dihydroxy-2-naphthoate = a demethylmenaquinol + diphosphate + CO2
Glossary: menaquinone = vitamin K2
Systematic name: all-trans-polyprenyl diphosphate:1,4-dihydroxy-2-naphthoate polyprenyltransferase
Comments: This enzyme catalyses a step in the synthesis of menaquinone, in which the prenyl chain synthesized by polyprenyl diphosphate synthase is transferred to 1,4-dihydroxy-2-naphthoate (DHNA). The bacterial enzyme is an inner membrane protein [1], with the C-terminus located in the periplasm [3]. It is highly specific for DHNA but not for a specific length of the prenyl chain [2].
References:
1. Shineberg, B. and Young, I.G. Biosynthesis of bacterial menaquinones: the membrane-associated 1,4-dihydroxy-2-naphthoate octaprenyltransferase of Escherichia coli. Biochemistry 15 (1976) 2754-2758. [PMID: 949474]
2. Saito, Y. and Ogura, K. Biosynthesis of menaquinones. Enzymatic prenylation of 1,4-dihydroxy-2-naphthoate by Micrococcus luteus membrane fractions. J. Biochem. 89 (1981) 1445-1452. [PMID: 7275947]
3. Suvarna, K., Stevenson, D., Meganathan, R. and Hudspeth, M.E. Menaquinone (vitamin K2) biosynthesis: localization and characterization of the menA gene from Escherichia coli. J. Bacteriol. 180 (1998) 2782-2787. [PMID: 9573170]
4. Daley, D.O., Rapp, M., Granseth, E., Melen, K., Drew, D. and von Heijne, G. Global topology analysis of the Escherichia coli inner membrane proteome. Science 308 (2005) 1321-1323. [PMID: 15919996]
[EC 2.5.1.74 created 2009]
EC 2.5.1.75
Accepted name: tRNA dimethylallyltransferase
Reaction: dimethyallyl diphosphate + tRNA = diphosphate + tRNA containing 6-dimethyallyladenosine
Other name(s): tRNA prenyltransferase, MiaA, transfer ribonucleate isopentenyltransferase (incorrect); Δ2-isopentenyl pyrophosphate:tRNA-Δ2-isopentenyl transferase (incorrect); Δ2-isopentenyl pyrophosphate:transfer ribonucleic acid Δ2-isopentenyltransferase (incorrect)
Systematic name: dimethyallyl-diphosphate: tRNA dimethylallyltransferase
Comments: Formerly known as tRNA isopentenyltransferase EC 2.5.1.8), but it is now known that dimethyallyl diphosphate, rather than isopentenyl diphosphate, is the substrate
References:
1. Leung, H.C., Chen, Y. and Winkler, M.E. Regulation of substrate recognition by the MiaA tRNA prenyltransferase modification enzyme of Escherichia coli K-12. J. Biol. Chem. 272 (1997) 13073-13083. [PMID: 9148919]
2. Soderberg, T. and Poulter, C.D. Escherichia coli dimethylallyl diphosphate:tRNA dimethylallyltransferase: essential elements for recognition of tRNA substrates within the anticodon stem-loop. Biochemistry 39 (2000) 6546-6553. [PMID: 10828971]
3. Moore, J.A., Mathis, J.R. and Poulter, C.D. Escherichia coli dimethylallyl diphosphate:tRNA dimethylallyltransferase: pre-steady-state kinetic studies. Biochim. Biophys. Acta 1479 (2000) 166-174. [PMID: 11004538]
[EC 2.5.1.75 created 1972 as EC 2.5.1.8, transferred 2009 to EC 2.5.1.75]
EC 2.5.1.76
Accepted name: cysteate synthase
Reaction: O-phospho-L-serine + sulfite = L-cysteate + phosphate
Systematic name: sulfite:O-phospho-L-serine sulfotransferase (phosphate-hydrolysing, L-cysteate-forming).
Comments: A pyridoxal-phosphate protein. It is highly specific for O-phospho-L-serine and sulfite. The reaction proceeds through a dehydroalanine (2-aminoacrylic acid) intermediate. The enzyme from Methanosarcina acetivorans is evolutionarily related to threonine synthase (EC 4.2.3.1), but the reaction is more similar to that of O-phosphoserine sulfhydrylase (EC 2.5.1.65).
References:
1. Graham, D.E., Taylor, S.M., Wolf, R.Z. and Namboori, S.C. Convergent evolution of coenzyme M biosynthesis in the Methanosarcinales: cysteate synthase evolved from an ancestral threonine synthase. Biochem. J. 424 (2009) 467-478. [PMID: 19761441]
[EC 2.5.1.76 created 2009]
*EC 2.7.1.119
Accepted name: hygromycin-B 7"-O-kinase
Reaction: ATP + hygromycin B = ADP + 7"-O-phosphohygromycin
For diagram click here
Other name(s): hygromycin B phosphotransferase; hygromycin-B kinase (ambiguous)
Systematic name: ATP:hygromycin-B 7"-O-phosphotransferase
Comments: Phosphorylates the antibiotics hygromycin B, 1-N-hygromycin B and destomycin, but not hygromycin B2, at the 7"-hydroxy group in the destomic acid ring.
Links to other databases:
BRENDA,
EXPASY,
KEGG,
CAS registry number: 88361-67-5
References:
1. Zalacain, M., Pardo, J.M. and Jiménez, A. Purification and characterization of a hygromycin B phosphotransferase from Streptomyces hygroscopicus. Eur. J. Biochem. 162 (1987) 419-422. [PMID: 3026811]
[EC 2.7.1.119 created 1989, modified 2009]
EC 2.7.1.163
Accepted name: hygromycin B 4-O-kinase
Reaction: ATP + hygromycin B = ADP + 4-O-phosphohygromycin B
Other name(s): hygromycin-B kinase (ambiguous)
Systematic name: ATP:hygromycin-B 4-O-phosphotransferase
Comments: Phosphorylates the antibiotic hygromycin B. Whereas the enzyme from Streptomyces hygroscopicus (EC 2.7.1.119; hygromycin-B 7"-O-kinase) catalyses the formation of 7"-O-phosphohygromycin B, this enzyme, found in Escherichia coli carrying a plasmid conferring resistance to hygromycin-B, forms 4-O-phosphohygromycin B.
References:
1. Rao, R.N., Allen, N.E., Hobbs, J.N., Jr., Alborn, W.E., Jr., Kirst, H.A. and Paschal, J.W. Genetic and enzymatic basis of hygromycin B resistance in Escherichia coli. Antimicrob. Agents Chemother. 24 (1983) 689-695. [PMID: 6318654]
[EC 2.7.1.163 created 2009]
EC 2.7.1.164
Accepted name: O-phosphoseryl-tRNASec kinase
Reaction: ATP + L-seryl-tRNASec = ADP + O-phospho-L-seryl-tRNASec
Other name(s): PSTK; phosphoseryl-tRNA[Ser]Sec kinase; phosphoseryl-tRNASec kinase
Systematic name: ATP:L-seryl-tRNASec O-phosphotransferase
Comments: In archaea and eukarya selenocysteine formation is achieved by a two-step process: O-phosphoseryl-tRNASec kinase (PSTK) phosphorylates the endogenous L-seryl-tRNASec to O-phospho-L-seryl-tRNASec, and then this misacylated amino acid-tRNA species is converted to L-selenocysteinyl-tRNASec by EC 2.9.1.2 (Sep-tRNA:Sec-tRNA synthase).
References:
1. Carlson, B.A., Xu, X.M., Kryukov, G.V., Rao, M., Berry, M.J., Gladyshev, V.N. and Hatfield, D.L. Identification and characterization of phosphoseryl-tRNA[Ser]Sec kinase. Proc. Natl. Acad. Sci. USA 101 (2004) 12848-12853. [PMID: 15317934]
2. Sherrer, R.L., O'Donoghue, P. and Soll, D. Characterization and evolutionary history of an archaeal kinase involved in selenocysteinyl-tRNA formation. Nucleic Acids Res. 36 (2008) 1187-1199. [PMID: 18174226]
3. Khangulov, S.V., Gladyshev, V.N., Dismukes, G.C. and Stadtman, T.C. Selenium-containing formate dehydrogenase H from Escherichia coli: a molybdopterin enzyme that catalyzes formate oxidation without oxygen transfer. Biochemistry 37 (1998) 3518-3528. [PMID: 9521673]
[EC 2.7.1.164 created 2009]
EC 2.9.1.2
Accepted name: O-phospho-L-seryl-tRNASec:L-selenocysteinyl-tRNA synthase
Reaction: O-phospho-L-seryl-tRNASec + selenophosphate = L-selenocysteinyl-tRNASec + phosphate
Other name(s): MMPSepSecS; SepSecS; SLA/LP; O-phosphoseryl-tRNA:selenocysteinyl-tRNA synthase; O-phospho-L-seryl-tRNA:L-selenocysteinyl-tRNA synthase
Systematic name: selenophosphate:O-phospho-L-seryl-tRNASec selenium transferase
Comments: A pyridoxal-phosphate protein [17142313]. In archaea and eukarya selenocysteine formation is achieved by a two-step process: EC 2.7.1.164 (O-phosphoseryl-tRNASec kinase) phosphorylates the endogenous L-seryl-tRNASec to O-phospho-L-seryl-tRNASec, and then this misacylated amino acid-tRNA species is converted to L-selenocysteinyl-tRNASec by Sep-tRNA:Sec-tRNA synthase.
References:
1. Palioura, S., Sherrer, R.L., Steitz, T.A., Soll, D. and Simonovic, M. The human SepSecS-tRNASec complex reveals the mechanism of selenocysteine formation. Science 325 (2009) 321-325. [PMID: 19608919]
2. Araiso, Y., Palioura, S., Ishitani, R., Sherrer, R.L., O'Donoghue, P., Yuan, J., Oshikane, H., Domae, N., Defranco, J., Soll, D. and Nureki, O. Structural insights into RNA-dependent eukaryal and archaeal selenocysteine formation. Nucleic Acids Res. 36 (2008) 1187-1199. [PMID: 18158303]
3. Aeby, E., Palioura, S., Pusnik, M., Marazzi, J., Lieberman, A., Ullu, E., Soll, D. and Schneider, A. The canonical pathway for selenocysteine insertion is dispensable in Trypanosomes. Proc. Natl. Acad. Sci. USA 106 (2009) 5088-5092. [PMID: 19279205]
[EC 2.9.1.2 created 2009]
EC 3.1.26.13
Accepted name: retroviral ribonuclease H
Reaction: Endohydrolysis of RNA in RNA/DNA hybrids. Three different cleavage modes:
1. sequence-specific internal cleavage of RNA [1-4]. Human immunodeficiency virus type 1 and Moloney murine leukemia virus enzymes prefer to cleave the RNA strand one nucleotide away from the RNA-DNA junction [5].
2. RNA 5'-end directed cleavage 13-19 nucleotides from the RNA end [6,7].
3. DNA 3'-end directed cleavage 15-20 nucleotides away from the primer terminus [8-10].
Other name(s): RT/RNase H; retroviral reverse transcriptase RNaseH; HIV RNase H
Comments: Comments: Retroviral reverse transcriptase is a multifunctional enzyme responsible for viral replication. To perform this task the enzyme combines two distinct activities. The polymerase domain (EC 2.7.7.49, RNA-directed DNA polymerase) occupies the N-terminal two-thirds of the reverse transcriptase whereas the ribonuclease H domain comprises the C-terminal remaining one-third [13,14].
The RNase H domain of Moloney murine leukemia virus and Human immunodeficiency virus display two metal binding sites [15-17]
References:
1. Schultz, S.J., Zhang, M. and Champoux, J.J. Recognition of internal cleavage sites by retroviral RNases H. J. Mol. Biol. 344 (2004) 635-652. [PMID: 15533434]
2. Sarafianos, S.G., Das, K., Tantillo, C., Clark, A.D., Jr., Ding, J., Whitcomb, J.M., Boyer, P.L., Hughes, S.H. and Arnold, E. Crystal structure of HIV-1 reverse transcriptase in complex with a polypurine tract RNA:DNA. EMBO J. 20 (2001) 1449-1461. [PMID: 11250910]
3. Rausch, J.W., Lener, D., Miller, J.T., Julias, J.G., Hughes, S.H. and Le Grice, S.F. Altering the RNase H primer grip of human immunodeficiency virus reverse transcriptase modifies cleavage specificity. Biochemistry 41 (2002) 4856-4865. [PMID: 11939780]
4. Brehm, J.H., Mellors, J.W. and Sluis-Cremer, N. Mechanism by which a glutamine to leucine substitution at residue 509 in the ribonuclease H domain of HIV-1 reverse transcriptase confers zidovudine resistance. Biochemistry 47 (2008) 14020-14027. [PMID: 19067547]
5. Schultz, S.J., Zhang, M., Kelleher, C.D. and Champoux, J.J. Analysis of plus-strand primer selection, removal, and reutilization by retroviral reverse transcriptases. J. Biol. Chem. 275 (2000) 32299-32309. [PMID: 10913435]
6. DeStefano, J.J., Mallaber, L.M., Fay, P.J. and Bambara, R.A. Determinants of the RNase H cleavage specificity of human immunodeficiency virus reverse transcriptase. Nucleic Acids Res. 21 (1993) 4330-4338. [PMID: 7692401]
7. Kati, W.M., Johnson, K.A., Jerva, L.F. and Anderson, K.S. Mechanism and fidelity of HIV reverse transcriptase. J. Biol. Chem. 267 (1992) 25988-25997. [PMID: 1281479]
8. Palaniappan, C., Fuentes, G.M., Rodriguez-Rodriguez, L., Fay, P.J. and Bambara, R.A. Helix structure and ends of RNA/DNA hybrids direct the cleavage specificity of HIV-1 reverse transcriptase RNase H. J. Biol. Chem. 271 (1996) 2063-2070. [PMID: 8567660]
9. Fu, T.B. and Taylor, J. When retroviral reverse transcriptases reach the end of their RNA templates. J. Virol. 66 (1992) 4271-4278. [PMID: 1376369]
10. Beilhartz, G.L., Wendeler, M., Baichoo, N., Rausch, J., Le Grice, S. and Gotte, M. HIV-1 reverse transcriptase can simultaneously engage its DNA/RNA substrate at both DNA polymerase and RNase H active sites: implications for RNase H inhibition. J. Mol. Biol. 388 (2009) 462-474. [PMID: 19289131]
11. Huang, H., Chopra, R., Verdine, G.L. and Harrison, S.C. Structure of a covalently trapped catalytic complex of HIV-1 reverse transcriptase: implications for drug resistance. Science 282 (1998) 1669-1675. [PMID: 9831551]
12. Krug, M.S. and Berger, S.L. Ribonuclease H activities associated with viral reverse transcriptases are endonucleases. Proc. Natl. Acad. Sci. USA 86 (1989) 3539-3543. [PMID: 2471188]
13. Champoux, J.J. and Schultz, S.J. Ribonuclease H: properties, substrate specificity and roles in retroviral reverse transcription. FEBS J. 276 (2009) 1506-1516. [PMID: 19228195]
14. Schultz, S.J. and Champoux, J.J. RNase H activity: structure, specificity, and function in reverse transcription. Virus Res 134 (2008) 86-103. [PMID: 18261820]
15. Goedken, E.R. and Marqusee, S. Metal binding and activation of the ribonuclease H domain from moloney murine leukemia virus. Protein Eng. 12 (1999) 975-980. [PMID: 10585503]
16. Davies, J.F., 2nd, Hostomska, Z., Hostomsky, Z., Jordan, S.R. and Matthews, D.A. Crystal structure of the ribonuclease H domain of HIV-1 reverse transcriptase. Science 252 (1991) 88-95. [PMID: 1707186]
17. Pari, K., Mueller, G.A., DeRose, E.F., Kirby, T.W. and London, R.E. Solution structure of the RNase H domain of the HIV-1 reverse transcriptase in the presence of magnesium. Biochemistry 42 (2003) 639-650. [PMID: 12534276]
[EC 3.1.26.13 created 2009]
EC 3.2.2.27
Accepted name: uracil-DNA glycosylase
Reaction: Hydrolyzes single-stranded DNA or mismatched double-stranded DNA and polynucleotides, releasing free uracil
Other name(s): UdgB (ambiguous); uracil-DNA N-glycosylase; UDG (ambiguous); uracil DNA glycohydrolase
Systematic name: uracil-DNA deoxyribohydrolase (uracil-releasing)
Comments: Uracil-DNA glycosylases are widespread enzymes that are found in all living organisms. EC 3.2.2.27 and double-stranded uracil-DNA glycosylase (EC 3.2.2.28) form a central part of the DNA-repair machinery since they initiate the DNA base-excision repair pathway by hydrolysing the N-glycosidic bond between uracil and the deoxyribose sugar thereby catalysing the removal of mis-incorporated uracil from DNA.
References:
1. Lee, M.S., Kim, G.A., Seo, M.S., Lee, J.H. and Kwon, S.T. Characterization of heat-labile uracil-DNA glycosylase from Psychrobacter sp. HJ147 and its application to the polymerase chain reaction. Biotechnol. Appl. Biochem. 52 (2009) 167-175. [PMID: 18412541]
2. Kim, G.A., Lee, M.S., Sun, Y., Lee, B.D., Lee, J.I., Lee, J.H. and Kwon, S.T. Characterization of cold-active uracil-DNA glycosylase from Bacillus sp. HJ171 and its use for contamination control in PCR. Appl. Microbiol. Biotechnol. 80 (2008) 785-794. [PMID: 18626641]
3. Parikh, S.S., Putnam, C.D. and Tainer, J.A. Lessons learned from structural results on uracil-DNA glycosylase. Mutat Res 460 (2000) 183-199. [PMID: 10946228]
4. Stivers, J.T. and Drohat, A.C. Uracil DNA glycosylase: insights from a master catalyst. Arch. Biochem. Biophys. 396 (2001) 1-9. [PMID: 11716455]
[EC 3.2.2.27 created 2009]
EC 3.2.2.28
Accepted name: double-stranded uracil-DNA glycosylase
Reaction: Specifically hydrolyzes mismatched double-stranded DNA and polynucleotides, releasing free uracil
Other name(s): Mug; double-strand uracil-DNA glycosylase; Dug; dsUDG; double-stranded DNA specific UDG; dsDNA specific UDG; UdgB (ambiguous); G:T/U mismatch-specific DNA glycosylase; UDG (ambiguous)
Systematic name: uracil-double-stranded DNA deoxyribohydrolase (uracil-releasing)
Comments: No activity on DNA containing a T/G mispair or single-stranded DNA containing either a site-specific uracil or 3,N4-ethenocytosine residue [2], significant role for double-stranded uracil-DNA glycosylase in mutation avoidance in non-dividing E. coli [3]. Uracil-DNA glycosylases are widespread enzymes that are found in all living organisms. Uracil-DNA glycosylase (EC 3.2.2.27) and EC 3.2.2.28 form a central part of the DNA-repair machinery since they initiate the DNA base-excision repair pathway by hydrolysing the N-glycosidic bond between uracil and the deoxyribose sugar thereby catalysing the removal of mis-incorporated uracil from DNA.
References:
1. Barrett, T.E., Scharer, O.D., Savva, R., Brown, T., Jiricny, J., Verdine, G.L. and Pearl, L.H. Crystal structure of a thwarted mismatch glycosylase DNA repair complex. EMBO J. 18 (1999) 6599-6609. [PMID: 10581234]
2. Sung, J.S. and Mosbaugh, D.W. Escherichia coli double-strand uracil-DNA glycosylase: involvement in uracil-mediated DNA base excision repair and stimulation of activity by endonuclease IV. Biochemistry 39 (2000) 10224-10235. [PMID: 10956012]
3. Mokkapati, S.K., Fernandez de Henestrosa, A.R. and Bhagwat, A.S. Escherichia coli DNA glycosylase Mug: a growth-regulated enzyme required for mutation avoidance in stationary-phase cells. Mol. Microbiol. 41 (2001) 1101-1111. [PMID: 11555290]
[EC 3.2.2.28 created 2009]
EC 3.2.2.29
Accepted name: thymine-DNA glycosylase
Reaction: Hydrolyzes mismatched double-stranded DNA and polynucleotides, releasing free thymine
Other name(s): mismatch-specific thymine-DNA glycosylase; mismatch-specific thymine-DNA N-glycosylase; hTDG; hsTDG; TDG; thymine DNA glycosylase; G/T glycosylase; uracil/thymine DNA glycosylase; T:G mismatch-specific thymidine-DNA glycosylase; G:T mismatch-specific thymine DNA-glycosylase
Systematic name: thymine-DNA deoxyribohydrolase (thymine-releasing)
Comments: Thymine-DNA glycosylase is part of the DNA-repair machinery. Thymine removal is fastest when it is from a G/T mismatch with a 5'-flanking C/G pair. The glycosylase removes uracil from G/U, C/U, and T/U base pairs faster than it removes thymine from G/T [3].
References:
1. Waters, T.R. and Swann, P.F. Thymine-DNA glycosylase and G to A transition mutations at CpG sites. Mutat Res 462 (2000) 137-147. [PMID: 10767625]
2. Neddermann, P. and Jiricny, J. The purification of a mismatch-specific thymine-DNA glycosylase from HeLa cells. J. Biol. Chem. 268 (1993) 21218-21224. [PMID: 8407958]
3. Waters, T.R. and Swann, P.F. Kinetics of the action of thymine DNA glycosylase. J. Biol. Chem. 273 (1998) 20007-20014. [PMID: 9685338]
[EC 3.2.2.29 created 2009]
EC 3.4.17.23
Accepted name: angiotensin-converting enzyme 2
Reaction: angiotensin II + H2O = angiotensin-(1-7) + L-phenylalanine
Other name(s): ACE-2; ACE2; hACE2; angiotensin converting enzyme 2; angiotensin converting enzyme-2; Tmem27
Comments: A transmembrane glycoprotein with an extracellular catalytic domain. Angiotensin-converting enzyme 2 functions as a carboxypeptidase, cleaving a single C-terminal residue from a distinct range of substrates [2]. Catalytic efficiency is 400-fold higher with angiotensin II (1-8) as a substrate than with angiotensin I (1-10). Angiotensin-converting enzyme 2 also efficiently hydrolyzes des-Arg9-bradykinin, but it does not hydrolyze bradykinin [1]. In peptidase family M2.
References:
1. Vickers, C., Hales, P., Kaushik, V., Dick, L., Gavin, J., Tang, J., Godbout, K., Parsons, T., Baronas, E., Hsieh, F., Acton, S., Patane, M., Nichols, A. and Tummino, P. Hydrolysis of biological peptides by human angiotensin-converting enzyme-related carboxypeptidase. J. Biol. Chem. 277 (2002) 14838-14843. [PMID: 11815627]
2. Lambert, D.W., Hooper, N.M. and Turner, A.J. Angiotensin-converting enzyme 2 and new insights into the renin-angiotensin system. Biochem. Pharmacol. 75 (2008) 781-786. [PMID: 17897633]
3. Towler, P., Staker, B., Prasad, S.G., Menon, S., Tang, J., Parsons, T., Ryan, D., Fisher, M., Williams, D., Dales, N.A., Patane, M.A. and Pantoliano, M.W. ACE2 X-ray structures reveal a large hinge-bending motion important for inhibitor binding and catalysis. J. Biol. Chem. 279 (2004) 17996-18007. [PMID: 14754895]
[EC 3.4.17.23 created 2009]
EC 3.4.22.69
Accepted name: SARS coronavirus main proteinase
Reaction: TSAVLQ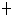 SGFRK-NH2 and SGVTFQGKFKK the two peptides corresponding to the two self-cleavage sites of the SARS 3C-like proteinase are the two most reactive peptide substrates. The enzyme exhibits a strong preference for substrates containing Gln at P1 position and Leu at P2 position.
SGFRK-NH2 and SGVTFQGKFKK the two peptides corresponding to the two self-cleavage sites of the SARS 3C-like proteinase are the two most reactive peptide substrates. The enzyme exhibits a strong preference for substrates containing Gln at P1 position and Leu at P2 position.
Other name(s): 3cLpro; 3C-like protease; coronavirus 3C-like protease; Mpro; SARS 3C-like protease; SARS coronavirus 3CL protease; SARS coronavirus main peptidase; SARS coronavirus main protease; SARS-CoV 3CLpro enzyme; SARS-CoV main protease; SARS-CoV Mpro; severe acute respiratory syndrome coronavirus main protease
Comments: SARS coronavirus main protease is the key enzyme in SARS coronavirus replicase polyprotein processing. In peptidase family C30.
References:
1. Goetz, D.H., Choe, Y., Hansell, E., Chen, Y.T., McDowell, M., Jonsson, C.B., Roush, W.R., McKerrow, J. and Craik, C.S. Substrate specificity profiling and identification of a new class of inhibitor for the major protease of the SARS coronavirus. Biochemistry 46 (2007) 8744-8752. [PMID: 17605471]
2. Fan, K., Wei, P., Feng, Q., Chen, S., Huang, C., Ma, L., Lai, B., Pei, J., Liu, Y., Chen, J. and Lai, L. Biosynthesis, purification, and substrate specificity of severe acute respiratory syndrome coronavirus 3C-like proteinase. J. Biol. Chem. 279 (2004) 1637-1642. [PMID: 14561748]
3. Akaji, K., Konno, H., Onozuka, M., Makino, A., Saito, H. and Nosaka, K. Evaluation of peptide-aldehyde inhibitors using R188I mutant of SARS 3CL protease as a proteolysis-resistant mutant. Bioorg. Med. Chem. 16:9400 (2008). [PMID: 18845442]
[EC 3.4.22.69 created 2009]
EC 3.4.22.70
Accepted name: sortase A
Reaction: The enzyme catalyzes a cell wall sorting reaction in which a surface protein with a sorting signal containing a LPXTG motif is cleaved between the Thr and Gly residue. The resulting threonine carboxyl end of the protein is covalently attached to a pentaglycine cross-bridge of peptidoglycan
Other name(s): SrtA; SrtA protein; SrtA sortase
Comments: In peptidase family C60.
References:
1. Ton-That, H., Liu, G., Mazmanian, S.K., Faull, K.F. and Schneewind, O. Purification and characterization of sortase, the transpeptidase that cleaves surface proteins of Staphylococcus aureus at the LPXTG motif. Proc. Natl. Acad. Sci. USA 96 (1999) 12424-12429. [PMID: 10535938]
2. Zong, Y., Bice, T.W., Ton-That, H., Schneewind, O. and Narayana, S.V. Crystal structures of Staphylococcus aureus sortase A and its substrate complex. J. Biol. Chem. 279 (2004) 31383-31389. [PMID: 15117963]
3. Race, P.R., Bentley, M.L., Melvin, J.A., Crow, A., Hughes, R.K., Smith, W.D., Sessions, R.B., Kehoe, M.A., McCafferty, D.G. and Banfield, M.J. Crystal structure of Streptococcus pyogenes sortase A: implications for sortase mechanism. J. Biol. Chem. 284 (2009) 6924-6933. [PMID: 19129180]
[EC 3.4.22.70 created 2009]
EC 3.4.22.71
Accepted name: sortase B
Reaction: The enzyme catalyzes a cell wall sorting reaction in which a surface protein with a sorting signal containing a NXTN motif is cleaved. The resulting threonine carboxyl end of the protein is covalently attached to a pentaglycine cross-bridge of peptidoglycan.
Other name(s): SrtB
Comments: In peptidase family C60.
References:
1. Zong, Y., Mazmanian, S.K., Schneewind, O. and Narayana, S.V. The structure of sortase B, a cysteine transpeptidase that tethers surface protein to the Staphylococcus aureus cell wall. Structure 12 (2004) 105-112. [PMID: 14725770]
2. Bierne, H., Garandeau, C., Pucciarelli, M.G., Sabet, C., Newton, S., Garcia-del Portillo, F., Cossart, P. and Charbit, A. Sortase B, a new class of sortase in Listeria monocytogenes. J. Bacteriol. 186 (2004) 1972-1982. [PMID: 15028680]
[EC 3.4.22.71 created 2009]
EC 3.4.23.50
Accepted name: human endogenous retrovirus K endopeptidase
Reaction: Processing at the authentic HIV-1 PR recognition site and release of the mature p17 matrix and the p24 capsid protein, as a result of the cleavage of the -SQNYPIVQ- cleavage site.
Other name(s): human endogenous retrovirus K10 endopeptidase; endogenous retrovirus HERV-K10 putative protease; human endogenous retrovirus K retropepsin; HERV K10 endopeptidase; HERV K10 retropepsin; HERV-K PR; HERV-K protease; HERV-K113 protease; human endogenous retrovirus K113 protease; human retrovirus K10 retropepsin
Comments: In peptidase family A2.
References:
1. Towler, E.M., Gulnik, S.V., Bhat, T.N., Xie, D., Gustschina, E., Sumpter, T.R., Robertson, N., Jones, C., Sauter, M., Mueller-Lantzsch, N., Debouck, C. and Erickson, J.W. Functional characterization of the protease of human endogenous retrovirus, K10: can it complement HIV-1 protease. Biochemistry 37 (1998) 17137-17144. [PMID: 9860826]
[EC 3.4.23.50 created 2009]
EC 3.4.23.51
Accepted name: HycI peptidase
Reaction: This enzyme specifically removes a 32-amino acid peptide from the C-terminus of the precursor of the large subunit of hydrogenase 3 in Escherichia coli by cleavage at the C-terminal side of Arg537.
Other name(s): HycI; HycE processing protein
Comments: The reaction requires nickel to be bound to the precursor of the large subunit of hydrogenase 3. The endopeptidase uses the metal in the large subunit of [NiFe]-hydrogenases as a recognition motif [1]. In peptidase family A31.
References:
1. Theodoratou, E., Paschos, A., Magalon, A., Fritsche, E., Huber, R. and Bock, A. Nickel serves as a substrate recognition motif for the endopeptidase involved in hydrogenase maturation. Eur. J. Biochem. 267 (2000) 1995-1999. [PMID: 10727938]
2. Yang, F., Hu, W., Xu, H., Li, C., Xia, B. and Jin, C. Solution structure and backbone dynamics of an endopeptidase HycI from Escherichia coli: implications for mechanism of the [NiFe] hydrogenase maturation. J. Biol. Chem. 282 (2007) 3856-3863. [PMID: 17150961]
[EC 3.4.23.51 created 2009]
EC 3.4.24.87
Accepted name: ADAMTS13 endopeptidase
Reaction: The enzyme cleaves the von Willebrand factor at bond Tyr842Met843 within the A2 domain, i.e. Tyr1605Met1606 in von Willebrand factor.
Other name(s): ADAMTS VWF cleaving metalloprotease; ADAMTS-13; ADAMTS13; vWF-cleaving protease; VWF-CP; vWF-degrading protease; Upshaw factor; von Willebrand factor cleaving protease; ADAMTS13 peptidase
Comments: In peptidase family M12.
References:
1. Fujikawa, K., Suzuki, H., McMullen, B. and Chung, D. Purification of human von Willebrand factor-cleaving protease and its identification as a new member of the metalloproteinase family. Blood 98 (2001) 1662-1666. [PMID: 11535495]
2. Dong, J.F., Moake, J.L., Nolasco, L., Bernardo, A., Arceneaux, W., Shrimpton, C.N., Schade, A.J., McIntire, L.V., Fujikawa, K. and Lopez, J.A. ADAMTS-13 rapidly cleaves newly secreted ultralarge von Willebrand factor multimers on the endothelial surface under flowing conditions. Blood 100 (2002) 4033-4039. [PMID: 12393397]
[EC 3.4.24.87 created 2009]
EC 3.4.25.2
Accepted name: HslU—HslV peptidase
Reaction: ATP-dependent cleavage of the cell division inhibitor SulA. The central and the C-terminal regions are preferentially cleaved. Major cleavage sites: Ala80Ser81, Ala150Ser151, Leu54Gln55, Ile163His164, Leu67Thr68, Leu49Leu50, Leu65Trp66.
Other name(s): ClpQ; ClpYQ; ClpYQ protease; HslUV; HslV-HslU; HslV peptidase; ATP-dependent HslV-HslU proteinase; caseinolytic protease X; caseinolytic proteinase X; ClpXP ATP-dependent protease; ClpXP protease; ClpXP serine proteinase; Escherichia coli ClpXP serine proteinase; HslUV protease; HslUV proteinase; HslVU protease; HslVU proteinase; protease ClpYQ; protease CodWX; protease HslVU; proteinase ClpYQ; proteinase HslUV
Comments: In peptidase family T1.
References:
1. Wang, J., Rho, S.H., Park, H.H. and Eom, S.H. Correction of X-ray intensities from an HslV-HslU co-crystal containing lattice-translocation defects. Acta Crystallogr. D Biol. Crystallogr. 61 (2005) 932-941. [PMID: 15983416]
2. Nishii, W. and Takahashi, K. Determination of the cleavage sites in SulA, a cell division inhibitor, by the ATP-dependent HslVU protease from Escherichia coli. FEBS Lett. 553 (2003) 351-354. [PMID: 14572649]
3. Ramachandran, R., Hartmann, C., Song, H.K., Huber, R. and Bochtler, M. Functional interactions of HslV (ClpQ) with the ATPase HslU (ClpY). Proc. Natl. Acad. Sci. USA 99 (2002) 7396-7401. [PMID: 12032294]
4. Yoo, S.J., Seol, J.H., Shin, D.H., Rohrwild, M., Kang, M.S., Tanaka, K., Goldberg, A.L. and Chung, C.H. Purification and characterization of the heat shock proteins HslV and HslU that form a new ATP-dependent protease in Escherichia coli. J. Biol. Chem. 271 (1996) 14035-14040. [PMID: 8662828]
[EC 3.4.25.2 created 2009]
EC 3.5.1.99
Accepted name: fatty acid amide hydrolase
Reaction: (1) anandamide + H2O = arachidonic acid + ethanolamine
(2) oleamide + H2O = oleic acid + NH3
Glossary: anandamide = (5Z,8Z,11Z,14Z)-N-(2-hydroxyethyl)icosa-5,8,11,14-tetraenamide
Other name(s): FAAH; oleamide hydrolase; anandamide amidohydrolase
Systematic name: fatty acylamide amidohydrolase
Comments: Integral membrane protein, the enzyme is responsible for the catabolism of neuromodulatory fatty acid amides, including anandamide and oleamide, occurs in mammalia.
References:
1. Boger, D.L., Fecik, R.A., Patterson, J.E., Miyauchi, H., Patricelli, M.P. and Cravatt, B.F. Fatty acid amide hydrolase substrate specificity. Bioorg. Med. Chem. Lett. 10 (2000) 2613-2616. [PMID: 11128635]
2. Patricelli, M.P., Lashuel, H.A., Giang, D.K., Kelly, J.W. and Cravatt, B.F. Comparative characterization of a wild type and transmembrane domain-deleted fatty acid amide hydrolase: identification of the transmembrane domain as a site for oligomerization. Biochemistry 37 (1998) 15177-15187. [PMID: 9790682]
3. Patricelli, M.P. and Cravatt, B.F. Characterization and manipulation of the acyl chain selectivity of fatty acid amide hydrolase. Biochemistry 40 (2001) 6107-6115. [PMID: 11352748]
[EC 3.5.1.99 created 2009]
EC 3.5.1.100
Accepted name: (R)-amidase
Reaction: (1) (R)-piperazine-2-carboxamide + H2O = (R)-piperazine-2-carboxylic acid + NH3
(2) β-alaninamide + H2O = β-alanine + NH3
Other name(s): R-stereospecific amidase; R-amidase
Systematic name: (R)-piperazine-2-carboxamide amidohydrolase
Comments: In addition (R)-piperidine-3-carboxamide is hydrolysed to (R)-piperidine-3-carboxylic acid and NH3, and (R)-piperazine-2-tert-butylcarboxamide is hydrolysed to (R)-piperazine-2-tert-butylcarboxylic acid and NH3 with lower activity. The enzyme does not act on the other amide substrates for EC 3.5.1.4 (amidase).
References:
1. Komeda, H., Harada, H., Washika, S., Sakamoto, T., Ueda, M. and Asano, Y. A novel R-stereoselective amidase from Pseudomonas sp. MCI3434 acting on piperazine-2-tert-butylcarboxamide. Eur. J. Biochem. 271 (2004) 1580-1590. [PMID: 15066183]
[EC 3.5.1.100 created 2009]
EC 3.5.1.101
Accepted name: L-proline amide hydrolase
Reaction: (1) (S)-piperidine-2-carboxamide + H2O = (S)-piperidine-2-carboxylic acid + NH3
(2) L-prolinamide + H2O = L-proline + NH3
Glossary: L-pipecolate = piperidine-2-carboxylate
Other name(s): S-stereoselective piperazine-2-tert-butylcarboxamide hydrolase; LaaA; L-amino acid amidase
Systematic name: (S)-piperidine-2-carboxamide amidohydrolase
References:
1. Komeda, H., Harada, H., Washika, S., Sakamoto, T., Ueda, M. and Asano, Y. S-stereoselective piperazine-2-tert-butylcarboxamide hydrolase from Pseudomonas azotoformans IAM 1603 is a novel L-amino acid amidase. Eur. J. Biochem. 271 (2004) 1465-1475. [PMID: 15066172]
[EC 3.5.1.101 created 2009]
EC 3.6.4.12
Accepted name: DNA helicase
Reaction: ATP + H2O = ADP + phosphate
Other name(s): 3' to 5' DNA helicase; 3'-5' DNA helicase; 3'-5' PfDH; 5' to 3' DNA helicase; AvDH1; BACH1 helicase; BcMCM; BLM protein; BRCA1-associated C-terminal helicase; CeWRN-1; Dbp9p; DmRECQ5; DNA helicase 120; DNA helicase A; DNA helicase E; DNA helicase II; DNA helicase III; DNA helicase RECQL5β; DNA helicase VI; dnaB; DnaB helicase E1; helicase HDH IV; Hel E; helicase DnaB; helicase domain of bacteriophage T7 gene 4 protein helicase; PcrA helicase; UvrD; hHcsA; Hmi1p; hPif1; MCM helicase; MCM protein; MER3 helicase; MER3 protein; MPH1; PcrA; PcrA helicase; PDH120; PfDH A; Pfh1p; PIF1
Systematic name: ATP phosphohydrolase (DNA helix unwinding)
Comments: DNA helicases utilize the energy from ATP hydrolysis to unwind double-stranded DNA. Some of them unwind duplex DNA with a 3' to 5' polarity [1,3,5,8], others show 5' to 3' polarity [10,11,12,13] or unwind DNA in both directions [14,15]. Some helicases unwind DNA as well as RNA [9,10]. May be identical with EC 3.6.4.13 (RNA helicase).
References:
1. Ozsoy, A.Z., Sekelsky, J.J. and Matson, S.W. Biochemical characterization of the small isoform of Drosophila melanogaster RECQ5 helicase. Nucleic Acids Res. 29 (2001) 2986-2993. [PMID: 11452023]
2. Tanner, J.A., Watt, R.M., Chai, Y.B., Lu, L.Y., Lin, M.C., Peiris, J.S., Poon, L.L., Kung, H.F. and Huang, J.D. The severe acute respiratory syndrome (SARS) coronavirus NTPase/helicase belongs to a distinct class of 5' to 3' viral helicases. J. Biol. Chem. 278 (2003) 39578-39582. [PMID: 12917423]
3. Nakagawa, T., Flores-Rozas, H. and Kolodner, R.D. The MER3 helicase involved in meiotic crossing over is stimulated by single-stranded DNA-binding proteins and unwinds DNA in the 3' to 5' direction. J. Biol. Chem. 276 (2001) 31487-31493. [PMID: 11376001]
4. Lee, C. and Seo, Y.S. Isolation and characterization of a processive DNA helicase from the fission yeast Schizosaccharomyces pombe that translocates in a 5'-to-3' direction. Biochem. J. 334 (1998) 377-386. [PMID: 9716495]
5. Phan, T.N., Ehtesham, N.Z., Tuteja, R. and Tuteja, N. A novel nuclear DNA helicase with high specific activity from Pisum sativum catalytically translocates in the 3'→5' direction. Eur. J. Biochem. 270 (2003) 1735-1745. [PMID: 12694186]
6. Bernstein, D.A., Zittel, M.C. and Keck, J.L. High-resolution structure of the E. coli RecQ helicase catalytic core. EMBO J. 22 (2003) 4910-4921. [PMID: 14517231]
7. Pike, A.C., Shrestha, B., Popuri, V., Burgess-Brown, N., Muzzolini, L., Costantini, S., Vindigni, A. and Gileadi, O. Structure of the human RECQ1 helicase reveals a putative strand-separation pin. Proc. Natl. Acad. Sci. USA 106:1039 (2009). [PMID: 19151156]
8. Curti, E., Smerdon, S.J. and Davis, E.O. Characterization of the helicase activity and substrate specificity of Mycobacterium tuberculosis UvrD. J. Bacteriol. 189 (2007) 1542-1555. [PMID: 17158674]
9. Frick, D.N. The hepatitis C virus NS3 protein: a model RNA helicase and potential drug target. Curr. Issues Mol. Biol. 9 (2007) 1-20. [PMID: 17263143]
10. Ivanov, K.A. and Ziebuhr, J. Human coronavirus 229E nonstructural protein 13: characterization of duplex-unwinding, nucleoside triphosphatase, and RNA 5'-triphosphatase activities. J. Virol. 78 (2004) 7833-7838. [PMID: 15220459]
11. Ivessa, A.S., Zhou, J.Q., Schulz, V.P., Monson, E.K. and Zakian, V.A. Saccharomyces Rrm3p, a 5' to 3' DNA helicase that promotes replication fork progression through telomeric and subtelomeric DNA. Genes Dev. 16 (2002) 1383-1396. [PMID: 12050116]
12. Zhou, J.Q., Qi, H., Schulz, V.P., Mateyak, M.K., Monson, E.K. and Zakian, V.A. Schizosaccharomyces pombe pfh1+ encodes an essential 5' to 3' DNA helicase that is a member of the PIF1 subfamily of DNA helicases. Mol. Biol. Cell 13 (2002) 2180-2191. [PMID: 12058079]
13. George, T., Wen, Q., Griffiths, R., Ganesh, A., Meuth, M. and Sanders, C.M. Human Pif1 helicase unwinds synthetic DNA structures resembling stalled DNA replication forks. Nucleic Acids Res. 37 (2009) 6491-6502. [PMID: 19700773]
14. Naqvi, A., Tinsley, E. and Khan, S.A. Purification and characterization of the PcrA helicase of Bacillus anthracis. J. Bacteriol. 185 (2003) 6633-6639. [PMID: 14594837]
15. Ruiz-Maso, J.A., Anand, S.P., Espinosa, M., Khan, S.A. and del Solar, G. Genetic and biochemical characterization of the Streptococcus pneumoniae PcrA helicase and its role in plasmid rolling circle replication. J. Bacteriol. 188 (2006) 7416-7425. [PMID: 16936036]
[EC 3.6.4.12 created 2009]
EC 3.6.4.13
Accepted name: RNA helicase
Reaction: ATP + H2O = ADP + phosphate
Other name(s): CSFV NS3 helicase; DBP2; DbpA; DDX17; DDX25; DDX3; DDX3X; DDX3Y; DDX4; DDX5; DEAD-box protein DED1; DEAD-box RNA helicase; DEAH-box protein 2; DEAH-box RNA helicase; DED1; Dex(H/D) RNA helicase; EhDEAD1; EhDEAD1 RNA helicase; eIF4A helicase; KOKV helicase; Mtr4p; nonstructural protein 3 helicase; NPH-II; RHA; RNA helicase A; RNA helicase DDX3; RNA helicase Hera; RNA-dependent ATPase; TGBp1 NTPase/helicase domain; VRH1; GRTH/DDX25
Systematic name: ATP phosphohydrolase (RNA helix unwinding)
Comments: RNA helicases utilize the energy from ATP hydrolysis to unwind RNA. Some of them unwind RNA with a 3' to 5' polarity [3], other show 5' to 3' polarity [8]. Some helicases unwind DNA as well as RNA [7,8]. May be identical with EC 3.6.4.12 (DNA helicase).
References:
1. Cordin, O., Tanner, N.K., Doere, M., Linder, P. and Banroques, J. The newly discovered Q motif of DEAD-box RNA helicases regulates RNA-binding and helicase activity. EMBO J. 23 (2004) 2478-2487. [PMID: 15201868]
2. Rodamilans, B. and Montoya, G. Expression, purification, crystallization and preliminary X-ray diffraction analysis of the DDX3 RNA helicase domain. Acta Crystallogr. Sect. F Struct. Biol. Cryst. Commun. 63 (2007) 283-286. [PMID: 17401195]
3. Lee, C.G. and Hurwitz, J. A new RNA helicase isolated from HeLa cells that catalytically translocates in the 3' to 5' direction. J. Biol. Chem. 267 (1992) 4398-4407. [PMID: 1537828]
4. Li, S.C., Chung, M.C. and Chen, C.S. Cloning and characterization of a DEAD box RNA helicase from the viable seedlings of aged mung bean. Plant Mol. Biol. 47 (2001) 761-770. [PMID: 11785937]
5. Wu, J., Bera, A.K., Kuhn, R.J. and Smith, J.L. Structure of the Flavivirus helicase: implications for catalytic activity, protein interactions, and proteolytic processing. J. Virol. 79 (2005) 10268-10277. [PMID: 16051820]
6. Gross, C.H. and Shuman, S. The nucleoside triphosphatase and helicase activities of vaccinia virus NPH-II are essential for virus replication. J. Virol. 72 (1998) 4729-4736. [PMID: 9573237]
7. Frick, D.N. The hepatitis C virus NS3 protein: a model RNA helicase and potential drug target. Curr. Issues Mol. Biol. 9 (2007) 1-20. [PMID: 17263143]
[EC 3.6.4.13 created 2009]
EC 3.7.1.11
Accepted name: cyclohexane-1,2-dione hydrolase
Reaction: cyclohexane-1,2-dione + H2O = 6-oxohexanoate
Systematic name: cyclohexane-1,2-dione acylhydrolase (decyclizing)
Comments: Highly specific; does not act on cyclohexanone or cyclohexane-1,3-dione as substrate.
References:
1. Harder, J. Anaerobic degradation of cyclohexane-1,2-diol by a new Azoarcus species. Arch. Microbiol. 168 (1997) 199-204.
2. Fraas, S., Steinbach, A.K., Tabbert, A., Harder, J., Ermler, U., Tittmann, K., Meyer, A. and Kroneck P.M.H. Cyclohexane-1,2-dione hydrolase: A new tool to degrade alicyclic compounds. J. Mol. Catalysis B: Enzymatic 61 (2009) 47-49.
[EC 3.7.1.11 created 2009]
EC 4.1.1.90
Accepted name: peptidyl-glutamate 4-carboxylase
Reaction: peptidyl-4-carboxyglutamate + 2,3-epoxyphylloquinone + H2O = peptidyl-glutamate + CO2 + O2 + phylloquinone
For diagram of reaction, click here
Other name(s): vitamin K-dependent carboxylase; γ-glutamyl carboxylase
Systematic name: peptidyl-glutamate 4-carboxylase (2-methyl-3-phytyl-1,4-naphthoquinone-epoxidizing)
Comments: The enzyme can use various vitamin-K derivatives, including menaquinone, but does not contain iron. In the reverse direction the mechanism appears to involve the generation of a strong base by oxygenation of vitamin K. It catalyses the post-translational modification of several proteins of the blood-clotting system. 9-12 glutamate residues are converted to 4-carboxyglutamate (Gla) in a specific domain of the target protein.
References:
1. Dowd, P., Hershline, R., Ham, S.W. and Naganathan, S. Vitamin K and energy transduction: a base strength amplification mechanism. Science 269 (1995) 1684-1691. [PMID: 7569894]
2. Furie, B., Bouchard, B.A. and Furie, B.C. Vitamin K-dependent biosynthesis of γ-carboxyglutamic acid. Blood 93 (1999) 1798-1808. [PMID: 10068650]
3. Rishavy, M.A., Hallgren, K.W., Yakubenko, A.V., Shtofman, R.L., Runge, K.W. and Berkner, K.L. Bronsted analysis reveals Lys218 as the carboxylase active site base that deprotonates vitamin K hydroquinone to initiate vitamin K-dependent protein carboxylation. Biochemistry 45 (2006) 13239-13248. [PMID: 17073445]
4. Silva, P.J. and Ramos, M.J. Reaction mechanism of the vitamin K-dependent glutamate carboxylase: a computational study. J. Phys. Chem. B 111 (2007) 12883-12887. [PMID: 17935315]
[EC 4.1.1.90 created 2009]
EC 4.1.99.13
Accepted name: (6-4)DNA photolyase
Reaction: (6-4) photoproduct (in DNA) = 2 pyrimidine residues (in DNA)
For diagram of reaction, click here.
Other name(s): DNA photolyase; H64PRH; NF-10; phr (6-4); PL-(6-4); OtCPF1; (6-4) PHR; At64PHR
Systematic name: (6-4) photoproduct pyrimidine-lyase
Comments: A flavoprotein (FAD). The overall repair reaction consists of two distinct steps, one of which is light-independent and the other one light-dependent. In the initial light-independent step, a 6-iminium ion is thought to be generated via proton transfer induced by two histidines highly conserved among the (6-4) photolyases. This intermediate spontaneously rearranges to form an oxetane intermediate by intramolecular nucleophilic attack. In the subsequent light-driven reaction, one electron is believed to be transferred from the fully reduced FAD cofactor (FADH-) to the oxetane intermediate thus forming a neutral FADH radical and an anionic oxetane radical, which spontaneously fractures. The excess electron is then back-transferred to the flavin radical restoring the fully reduced flavin cofactor and a pair of pyrimidine bases [2].
References:
1. Hitomi, K., DiTacchio, L., Arvai, A.S., Yamamoto, J., Kim, S.T., Todo, T., Tainer, J.A., Iwai, S., Panda, S. and Getzoff, E.D. Functional motifs in the (6-4) photolyase crystal structure make a comparative framework for DNA repair photolyases and clock cryptochromes. Proc. Natl. Acad. Sci. USA 106 (2009) 6962-6967. [PMID: 19359474]
2. Schleicher, E., Hitomi, K., Kay, C.W., Getzoff, E.D., Todo, T. and Weber, S. Electron nuclear double resonance differentiates complementary roles for active site histidines in (6-4) photolyase. J. Biol. Chem. 282 (2007) 4738-4747. [PMID: 17164245]
[EC 4.1.99.13 created 2009]
EC 4.1.99.14
Accepted name: R-specific spore photoproduct lyase
Reaction: (5R)-5,6-dihydro-5-(thymidin-7-yl)thymidine (in double-helical DNA) + S-adenosyl-L-methionine = thymidylyl-(3'→5')-thymidylate (in double-helical DNA) + 5'-deoxyadenosine + L-methionine
Other name(s): SPL
Systematic name: R-specific spore photoproduct pyrimidine-lyase
Comments: The enzyme utilizes a [4Fe-4S] cluster and S-adenosyl-L-methionine as essential cofactors in spore photoproduct repair of double-helical DNA. The enzyme from Clostridium is specific for the (5R)-isomer of the methylene-bridged thymine dimer [1]. For the enzyme from Bacillus subtilis a (5S)-specificity was demonstrated with single-stranded DNA and synthetic substrates (cf. EC 4.1.99.15 [S-specific spore photoproduct lyase]).
References:
1. Chandra, T., Silver, S.C., Zilinskas, E., Shepard, E.M., Broderick, W.E. and Broderick, J.B. Spore photoproduct lyase catalyzes specific repair of the 5R but not the 5S spore photoproduct. J. Am. Chem. Soc. 131 (2009) 2420-2421. [PMID: 19178276]
[EC 4.1.99.14 created 2009]
EC 4.1.99.15
Accepted name: S-specific spore photoproduct lyase
Reaction: (5S)-5,6-dihydro-5-(thymidin-7-yl)thymidine (in DNA) + S-adenosyl-L-methionine = thymidylyl-(3'→5')-thymidylate (in DNA) + 5'-deoxyadenosine + L-methionine
For diagram click here
Other name(s): SAM; SP lyase; SPL; SplB; SplG
Systematic name: S-specific spore photoproduct pyrimidine-lyase
Comments: An iron-sulfur protein (contains one [4Fe-4S]2+ per enzyme monomer). The 5'-deoxy-adenosine radical formed after electron transfer from the [4Fe-4S] cluster to the S-adenosyl-L-methionine, initiates the repair by abstracting the C-6 hydrogen of the spore photoproduct lesion. The C-C bond linking the two pyrimidines undergoes fragmentation to give an allyl-type radical. The thermodynamically problematic last step is the transfer of the hydrogen atom back from the 5'-deoxyadenosine to the thymine monomer radical, which completes the repair process [3]. The enzyme from Bacillus subtilis is specific for the (5S)-configured spore photoproduct [4]. For the enzyme from Clostridium acetobutylicum a (5R)-specificity was demonstrated with spore photoproduct in double-helical DNA (cf. EC 4.1.99.14 [R-specific spore photoproduct lyase])
References:
1. Chandor, A., Berteau, O., Douki, T., Gasparutto, D., Sanakis, Y., Ollagnier-de-Choudens, S., Atta, M. and Fontecave, M. Dinucleotide spore photoproduct, a minimal substrate of the DNA repair spore photoproduct lyase enzyme from Bacillus subtilis. J. Biol. Chem. 281 (2006) 26922-26931. [PMID: 16829676]
2. Pieck, J.C., Hennecke, U., Pierik, A.J., Friedel, M.G. and Carell, T. Characterization of a new thermophilic spore photoproduct lyase from Geobacillus stearothermophilus (SplG) with defined lesion containing DNA substrates. J. Biol. Chem. 281 (2006) 36317-36326. [PMID: 16968710]
3. Mehl, R.A. and Begley, T.P. Mechanistic studies on the repair of a novel DNA photolesion: the spore photoproduct. Org Lett 1 (1999) 1065-1066. [PMID: 10825958]
4. Friedel, M.G., Berteau, O., Pieck, J.C., Atta, M., Ollagnier-de-Choudens, S., Fontecave, M. and Carell, T. The spore photoproduct lyase repairs the 5S- and not the 5R-configured spore photoproduct DNA lesion. Chem. Commun. (Camb.) (2006) 445-447. [PMID: 16493831]
[EC 4.1.99.15 created 2009]
EC 4.2.1.116
Accepted name: 3-hydroxypropionyl-CoA dehydratase
Reaction: 3-hydroxypropanoyl-CoA = acrylyl-CoA + H2O
Glossary: acrylyl-CoA = acryloyl-CoA
3-hydroxypropionyl-CoA = 3-hydroxypropanoyl-CoA
Systematic name: 3-hydroxypropionyl-CoA hydro-lyase
Comments: Catalyses a step in the 3-hydroxypropionate/4-hydroxybutyrate cycle, an autotrophic CO2 fixation pathway found in some thermoacidophilic archaea [1]. The enzyme from Metallosphaera sedula acts nearly equally as well on (S)-3-hydroxybutanoyl-CoA but not (R)-3-hydroxybutanoyl-CoA [2].
References:
1. Berg, I.A., Kockelkorn, D., Buckel, W. and Fuchs, G. A 3-hydroxypropionate/4-hydroxybutyrate autotrophic carbon dioxide assimilation pathway in Archaea. Science 318 (2007) 1782-1786. [PMID: 18079405]
2. Teufel, R., Kung, J.W., Kockelkorn, D., Alber, B.E. and Fuchs, G. 3-hydroxypropionyl-coenzyme A dehydratase and acryloyl-coenzyme A reductase, enzymes of the autotrophic 3-hydroxypropionate/4-hydroxybutyrate cycle in the Sulfolobales. J. Bacteriol. 191 (2009) 4572-4581. [PMID: 19429610]
[EC 4.2.1.116 created 2009]
EC 4.2.1.117
Accepted name: 2-methylcitrate dehydratase (2-methyl-trans-aconitate forming)
Reaction: (2S,3S)-2-methylcitrate = 2-methyl-trans-aconitate + H2O
Glossary: (2S,3S)-2-methylcitrate = (2S,3S)-2-hydroxybutane-1,2,3-tricarboxylate
Systematic name: (2S,3S)-2-hydroxybutane-1,2,3-tricarboxylate hydro-lyase (2-methyl-trans-aconitate forming)
Comments: Catalyses the dehydration of (2S,3S)-2-methylcitrate, forming the trans isomer of 2-methyl-aconitate (unlike EC 4.2.1.79, which forms only the cis isomer). Part of a propionate degradation pathway. The enzyme from Shewanella oneidensis can also accept citrate and cis-aconitate, but activity with (2S,3S)-2-methylcitrate was approximately 2.5-fold higher. 2-methylisocitrate and isocitrate were not substrates [1]. An iron-sulfur protein.
References:
1. Grimek, T.L. and Escalante-Semerena, J.C. The acnD genes of Shewenella oneidensis and Vibrio cholerae encode a new Fe/S-dependent 2-methylcitrate dehydratase enzyme that requires prpF function in vivo. J. Bacteriol. 186 (2004) 454-462. [PMID: 14702315]
[EC 4.2.1.117 created 2009]
EC 4.2.1.118
Accepted name: 3-dehydroshikimate dehydratase
Reaction: 3-dehydro-shikimate = protocatechuate + H2O
Systematic name: 3-dehydroshikimate hydro-lyase
Comments: Catalyses an early step in the biosynthesis of petrobactin, a siderophore produced by many bacteria, including the human pathogen Bacillus anthracis. Requires divalent ions, with a preference for Mn2+.
References:
1. Fox, D.T., Hotta, K., Kim, C.Y. and Koppisch, A.T. The missing link in petrobactin biosynthesis: asbF encodes a (–)-3-dehydroshikimate dehydratase. Biochemistry 47 (2008) 12251-12253. [PMID: 18975921]
2. Pfleger, B.F., Kim, Y., Nusca, T.D., Maltseva, N., Lee, J.Y., Rath, C.M., Scaglione, J.B., Janes, B.K., Anderson, E.C., Bergman, N.H., Hanna, P.C., Joachimiak, A. and Sherman, D.H. Structural and functional analysis of AsbF: origin of the stealth 3,4-dihydroxybenzoic acid subunit for petrobactin biosynthesis. Proc. Natl. Acad. Sci. USA 105 (2008) 17133-17138. [PMID: 18955706]
[EC 4.2.1.118 created 2009]
EC 4.2.1.119
Accepted name: enoyl-CoA hydratase 2
Reaction: (3R)-3-hydroxyacyl-CoA = (2E)-2-enoyl-CoA + H2O
Other name(s): 2-enoyl-CoA hydratase 2; AtECH2; ECH2; MaoC; MFE-2; PhaJAc; D-3-hydroxyacyl-CoA hydro-lyase; D-specific 2-trans-enoyl-CoA hydratase
Systematic name: (3R)-3-hydroxyacyl-CoA hydro-lyase
Comments: This enzyme catalyses a hydration step in peroxisomal β-oxidation. The human multifunctional enzyme type 2 (MFE-2) is a 79000 Da enzyme composed of three functional units: (3R)-hydroxyacyl-CoA dehydrogenase, 2-enoyl-CoA hydratase 2 and sterol carrier protein 2-like units [1]. The enzymes from Aeromonas caviae [4] and Arabidopsis thaliana [5] are monofunctional enzymes. 2-Enoyl-CoA hydratase 3 from Candida tropicalis is a part from multifunctional enzyme type 2 [3].
References:
1. Koski, K.M., Haapalainen, A.M., Hiltunen, J.K. and Glumoff, T. Crystal structure of 2-enoyl-CoA hydratase 2 from human peroxisomal multifunctional enzyme type 2. J. Mol. Biol. 345 (2005) 1157-1169. [PMID: 15644212]
2. Fukui, T., Shiomi, N. and Doi, Y. Expression and characterization of (R)-specific enoyl coenzyme A hydratase involved in polyhydroxyalkanoate biosynthesis by Aeromonas caviae. J. Bacteriol. 180 (1998) 667-673. [PMID: 9457873]
3. Koski, M.K., Haapalainen, A.M., Hiltunen, J.K. and Glumoff, T. Crystallization and preliminary crystallographic data of 2-enoyl-CoA hydratase 2 domain of Candida tropicalis peroxisomal multifunctional enzyme type 2. Acta Crystallogr. D Biol. Crystallogr. 59 (2003) 1302-1305. [PMID: 12832794]
4. Hisano, T., Fukui, T., Iwata, T. and Doi, Y. Crystallization and preliminary X-ray analysis of (R)-specific enoyl-CoA hydratase from Aeromonas caviae involved in polyhydroxyalkanoate biosynthesis. Acta Crystallogr. D Biol. Crystallogr. 57 (2001) 145-147. [PMID: 11134939]
5. Goepfert, S., Hiltunen, J.K. and Poirier, Y. Identification and functional characterization of a monofunctional peroxisomal enoyl-CoA hydratase 2 that participates in the degradation of even cis-unsaturated fatty acids in Arabidopsis thaliana. J. Biol. Chem. 281 (2006) 35894-35903. [PMID: 16982622]
6. Engeland, K. and Kindl, H. Evidence for a peroxisomal fatty acid β-oxidation involving D-3-hydroxyacyl-CoAs. Characterization of two forms of hydro-lyase that convert D-(-)-3-hydroxyacyl-CoA into 2-trans-enoyl-CoA. Eur. J. Biochem. 200 (1991) 171-178. [PMID: 1879422]
[EC 4.2.1.119 created 2009]
EC 4.2.1.120
Accepted name: 4-hydroxybutanoyl-CoA dehydratase
Reaction: 4-hydroxybutanoyl-CoA = but-3-enoyl-CoA + H2O
Glossary: 4-hydroxybutanoyl-CoA = 4-hydroxybutyryl-CoA
but-3-enoyl-CoA = vinylacetyl-CoA
Systematic name: 4-hydroxybutanoyl-CoA hydro-lyase
Comments: Contains FAD and a [3Fe-4S] iron-sulfur cluster. The enzyme is often present as a bifunctional enzyme, catalysing the dehydration of 4-hydroxybutanoyl-CoA to but-3-enoyl-CoA followed by isomerization of the later to crotonyl-CoA (EC 5.3.3.3). The enzyme has been characterized from several microorganisms, including Clostridium kluyveri, where it participates in succinate fermentation [1,2], Clostridium aminobutyricum, where it participates in 4-aminobutyrate degradation [3,4], and Metallosphaera sedula, where it participates in the 3-hydroxypropionate/4-hydroxybutyrate cycle, an autotrophic CO2 fixation pathway found in some thermoacidophilic archaea [5].
References:
1. Bartsch, R.G. and Barker, H.A. A vinylacetyl isomerase from Clostridium kluyveri. Arch. Biochem. Biophys. 92 (1961) 122-132. [PMID: 13687513]
2. Scherf, U., Sohling, B., Gottschalk, G., Linder, D. and Buckel, W. Succinate-ethanol fermentation in Clostridium kluyveri: purification and characterisation of 4-hydroxybutyryl-CoA dehydratase/vinylacetyl-CoA Δ3-Δ2-isomerase. Arch. Microbiol. 161 (1994) 239-245. [PMID: 8161284]
3. Scherf, U. and Buckel, W. Purification and properties of an iron-sulfur and FAD-containing 4-hydroxybutyryl-CoA dehydratase/vinylacetyl-CoA Δ3-Δ2-isomerase from Clostridium aminobutyricum. Eur. J. Biochem. 215 (1993) 421-429. [PMID: 8344309]
4. Muh, U., Cinkaya, I., Albracht, S.P. and Buckel, W. 4-Hydroxybutyryl-CoA dehydratase from Clostridium aminobutyricum: characterization of FAD and iron-sulfur clusters involved in an overall non-redox reaction. Biochemistry 35 (1996) 11710-11718. [PMID: 8794752]
5. Berg, I.A., Kockelkorn, D., Buckel, W. and Fuchs, G. A 3-hydroxypropionate/4-hydroxybutyrate autotrophic carbon dioxide assimilation pathway in Archaea. Science 318 (2007) 1782-1786. [PMID: 18079405]
[EC 4.2.1.120 created 2009]
EC 4.2.3.45
Accepted name: phyllocladan-16α-ol synthase
Reaction: (+)-copalyl diphosphate + H2O = phyllocladan-16α-ol + diphosphate
For diagram of reaction click here and for mechanism click here
Other name(s): PaDC1
Systematic name: (+)-copalyl-diphosphate diphosphate-lyase (phyllocladan-16α-ol-forming)
Comments: The adjacent gene PaDC2 codes EC 5.5.1.12 copalyl diphosphate synthase.
References:
1. Toyomasu, T., Niida, R., Kenmoku, H., Kanno, Y., Miura, S., Nakano, C., Shiono, Y., Mitsuhashi, W., Toshima, H., Oikawa, H., Hoshino, T., Dairi, T., Kato, N. and Sassa, T. Identification of diterpene biosynthetic gene clusters and functional analysis of labdane-related diterpene cyclases in Phomopsis amygdali. Biosci. Biotechnol. Biochem. 72 (2008) 1038-1047. [PMID: 18391465]
[EC 4.2.3.45 created 2009]
EC 4.99.1.8
Accepted name: heme ligase
Reaction: 2 ferriprotoporphyrin IX = β-hematin
Other name(s): heme detoxification protein; HDP; hemozoin synthase
Systematic name: Fe3+:ferriprotoporphyrin IX ligase (β-hematin-forming)
Comments: This heme detoxifying enzyme is found in Plasmodium parasites and converts toxic heme to crystalline hemozoin. These organisms lack the mammalian heme oxygenase for elimination of heme.
References:
1. Jani, D., Nagarkatti, R., Beatty, W., Angel, R., Slebodnick, C., Andersen, J., Kumar, S. and Rathore, D. HDP-a novel heme detoxification protein from the malaria parasite. PLoS Pathog. 4 (2008) e1000053. [PMID: 18437218]
[EC 4.99.1.8 created 2009]
EC 6.1.1.27
Accepted name: O-phospho-L-serine—tRNA ligase
Reaction: ATP + O-phospho-L-serine + tRNACys = AMP + diphosphate + O-phospho-L-seryl-tRNACys
Other name(s): non-canonical O-phosphoseryl-tRNA synthetase; SepRS; O-phosphoseryl-tRNA ligase
Systematic name: O-phospho-L-serine:tRNACys ligase (AMP-forming)
Comments: In organisms like Archaeoglobus fulgidus lacking EC 6.1.1.16 (cysteine—tRNA ligase) for the direct Cys-tRNACys formation, Cys-tRNACys is produced by an indirect pathway, in which EC 6.1.1.27 ligates O-phosphoserine to tRNACys, and EC 2.5.1.73 (O-phospho-L-seryl-tRNA: Cys-tRNA synthase) converts the produced O-phospho-L-seryl-tRNACys to Cys-tRNACys. The SepRS/SepCysS pathway is the sole route for cysteine biosynthesis in the organism [1]. Methanosarcina mazei can use both pathways, the direct route using EC 6.1.1.16 (cysteine—tRNA ligase) and the indirect pathway with EC 6.1.1.27 and EC 2.5.1.73 (O-phospho-L-seryl-tRNA: Cys-tRNA synthase) [2].
References:
1. Fukunaga, R. and Yokoyama, S. Structural insights into the first step of RNA-dependent cysteine biosynthesis in archaea. Nat. Struct. Mol. Biol. 14 (2007) 272-279. [PMID: 17351629]
2. Hauenstein, S.I. and Perona, J.J. Redundant synthesis of cysteinyl-tRNACys in Methanosarcina mazei. J. Biol. Chem. 283 (2008) 22007-22017. [PMID: 18559341]
[EC 6.1.1.27 created 2009]
EC 6.2.1.36
Accepted name: 3-hydroxypropionyl-CoA synthase
Reaction: 3-hydroxypropionate + ATP + coenzyme A = 3-hydroxypropionyl-CoA + AMP + diphosphate
Other name(s): 3-hydroxypropionyl-CoA synthetase (AMP-forming); 3-hydroxypropionate—CoA ligase
Systematic name: hydroxypropionate:CoA ligase (AMP-forming)
Comments: Catalyses a step in the 3-hydroxypropionate/4-hydroxybutyrate cycle, an autotrophic CO2 fixation pathway found in some thermoacidophilic archaea [1,2].
The enzymes from Metallosphaera sedula and Sulfolobus tokodaii can also use propionate, acrylate, acetate, and butanoate as substrates [2], and are thus different from EC 6.2.1.17 (propionate—CoA ligase), which does not accept acetate or butanoate.
References:
1. Berg, I.A., Kockelkorn, D., Buckel, W. and Fuchs, G. A 3-hydroxypropionate/4-hydroxybutyrate autotrophic carbon dioxide assimilation pathway in Archaea. Science 318 (2007) 1782-1786. [PMID: 18079405]
2. Alber, B.E., Kung, J.W. and Fuchs, G. 3-Hydroxypropionyl-coenzyme A synthetase from Metallosphaera sedula, an enzyme involved in autotrophic CO2 fixation. J. Bacteriol. 190 (2008) 1383-1389. [PMID: 18165310]
[EC 6.2.1.36 created 2009]
EC 6.3.1.13
Accepted name: L-cysteine:1D-myo-inositol 2-amino-2-deoxy-α-D-glucopyranoside ligase
Reaction: 1-O-(2-amino-2-deoxy-α-D-glucopyranosyl)-1D-myo-inositol + L-cysteine + ATP = 1-O-[2-(L-cysteinamido)-2-deoxy-α-D-glucopyranosyl]-1D-myo-inositol + AMP + diphosphate
Other name(s): MshC; MshC ligase; Cys:GlcN-Ins ligase; mycothiol ligase
Systematic name: L-cysteine:1-O-(2-amino-2-deoxy-α-D-glucopyranosyl)-1D-myo-inositol ligase (AMP-forming)
Comments: This enzyme is a key enzyme in the biosynthesis of mycothiol, a small molecular weight thiol found in Mycobacteria spp. and other actinomycetes. Mycothiol plays a fundamental role in these organisms by helping to provide protection from the effects of reactive oxygen species and electrophiles, including many antibiotics. The enzyme may represent a novel target for new classes of antituberculars [2]
References:
1. Fan, F., Luxenburger, A., Painter, G.F. and Blanchard, J.S. Steady-state and pre-steady-state kinetic analysis of Mycobacterium smegmatis cysteine ligase (MshC). Biochemistry 46 (2007) 11421-11429. [PMID: 17848100]
2. Gutierrez-Lugo, M.T., Newton, G.L., Fahey, R.C. and Bewley, C.A. Cloning, expression and rapid purification of active recombinant mycothiol ligase as B1 immunoglobulin binding domain of streptococcal protein G, glutathione-S-transferase and maltose binding protein fusion proteins in Mycobacterium smegmatis. Protein Expr. Purif. 50 (2006) 128-136. [PMID: 16908186]
3. Tremblay, L.W., Fan, F., Vetting, M.W. and Blanchard, J.S. The 1.6 Å crystal structure of Mycobacterium smegmatis MshC: the penultimate enzyme in the mycothiol biosynthetic pathway. Biochemistry 47 (2008) 13326-13335. [PMID: 19053270]
[EC 6.3.1.13 created 2009]